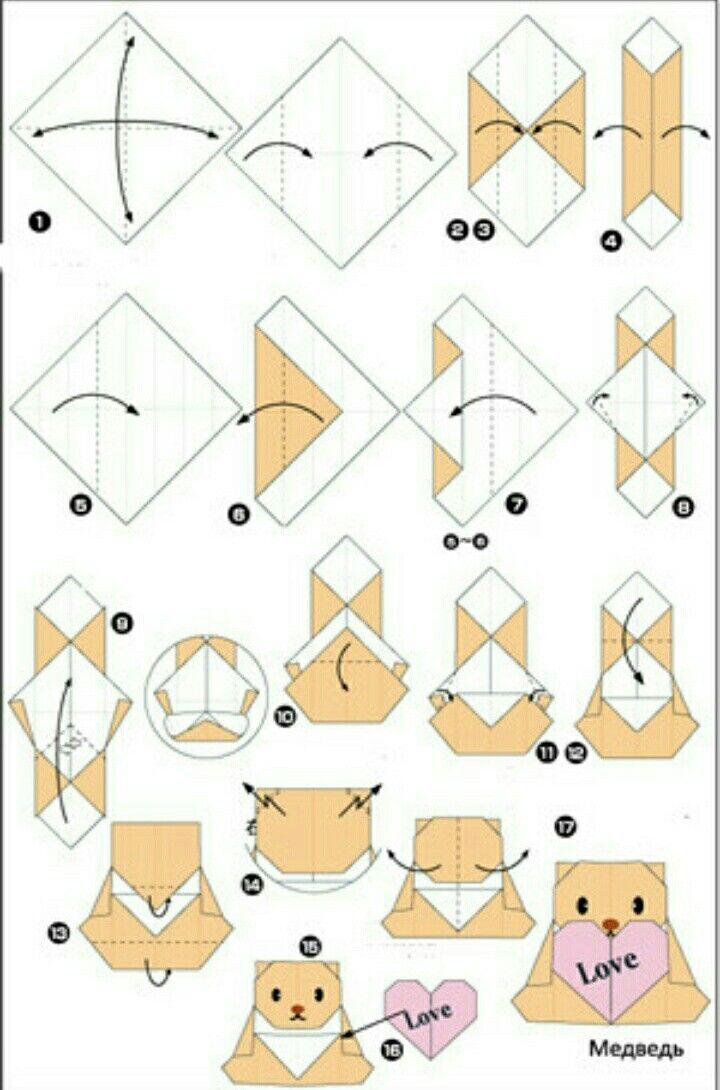
Узнаем как изготовить модульное оригами сердце?
Хотите украсить свою квартиру к празднику, но не знаете, чем? Сделайте модульное сердце. Оригами – это довольно простая техника, которая открывает множество возможностей. Из бумаги делают красивые изделия, которыми можно декорировать стены и дверные или оконные проемы. Инструкции по изготовлению ищите ниже.
Сложное оригами
Изготовить такую поделку будет несложно, если придерживаться схемы. Моделировать оригами «сердце» следует из прямоугольника. Располагаем его длинной стороной к себе. Теперь следует сделать конструктивные заломы. Сгибаем заготовку треугольником, как показано на схеме. Сначала следует проложить загибы сверху, а затем снизу. Приступаем к моделированию. Создаем из правой части треугольник путем складывания его по конструктивным линиям. Ту же самую операцию нужно проделать с противоположной стороной заготовки. Хотя можно вести работу и по частям. Сначала сложить правую сторону сердца, а затем левую. Края правого треугольника нужно притянуть к центру. Теперь следует разложить их в плоский треугольник. Подробную инструкцию этого процесса можно посмотреть на схеме. Ту же операцию нужно повторить с противоположной частью изделия. Теперь следует загнуть все острые части назад. Сердце готово. Из таких заготовок можно собрать гирлянду.
Края правого треугольника нужно притянуть к центру. Теперь следует разложить их в плоский треугольник. Подробную инструкцию этого процесса можно посмотреть на схеме. Ту же операцию нужно повторить с противоположной частью изделия. Теперь следует загнуть все острые части назад. Сердце готово. Из таких заготовок можно собрать гирлянду.
Сердце из модулей
Такой поделкой можно украсить свой дом к празднику всех влюбленных или к тематической вечеринке, например, для двойного свидания. Модульное объемное «сердце» (оригами) изготавливается из конусов. Сначала нужно сделать заготовки. Прямоугольник красной цветной бумаги нужно завернуть в конус и закрепить его в этом положении с помощью клея или же горячего пистолета. Чтобы собрать модульное оригами «сердце», потребуется основа. Вырезаем ее из картона. Теперь следует приклеить к нему заготовки. Начинать этот процесс нужно с наружного края и продвигаться в центр. Собираем один ряд, а затем вплотную к нему прикрепляем следующий. Удобнее всего это делать клеевым пистолетом.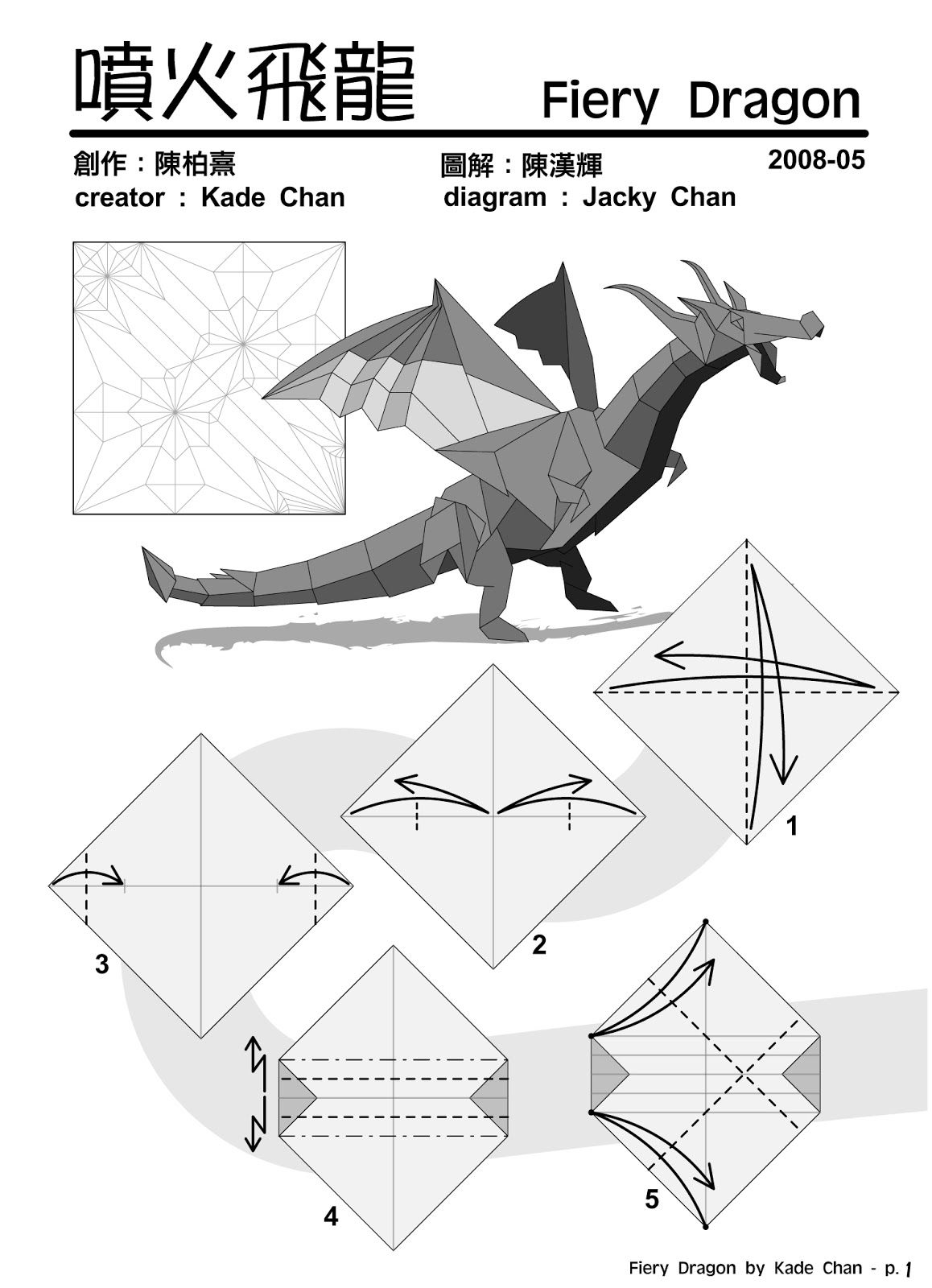 Когда весь объем заготовки заполнен, поделку можно считать законченной.
Когда весь объем заготовки заполнен, поделку можно считать законченной.
Простая поделка
Модульное оригами «сердце» можно собрать из полосок цветной бумаги. Эта техника называется квиллинг. Чаще всего такими изделиями украшают открытки и шкатулки. Но никто не запрещает выходить за эти рамки и создавать поделки, которые будут являться декором помещения. Из таких сердец можно сделать гирлянду или же нанизать их на нитку и собрать подвеску. Если у вас хватит сил и терпения, то можно сделать даже шторку, которую следует разместить или в дверном проеме, или же повесить в гостиной для зонирования помещения.
Как же собрать модульное оригами сердце? Вырезаем две тонкие полоски. Теперь следует склеить их снизу. Верхние края нужно согнуть сердечком и зафиксировать в таком положении. Внутренность заготовки тоже можно заполнить полосками, загибая их в различных направлениях и приклеивая к стенкам основы.
Бумажное сердце
Такую поделку тоже можно использовать для декора открытки или же для украшения помещения. Схема модульного «сердца» (оригами) приведена выше. Следует вырезать заготовки из двухсторонней цветной бумаги. Теперь нужно приклеить их друг к другу, фиксируя каждый слой в центре предыдущего.
Схема модульного «сердца» (оригами) приведена выше. Следует вырезать заготовки из двухсторонней цветной бумаги. Теперь нужно приклеить их друг к другу, фиксируя каждый слой в центре предыдущего.
Сердце можно собирать как из одноцветных заготовок, так и делать из разноцветных модулей.
Новые работы в технике Оригами модульное / Мастерклассы Блоги
Создание красивых фигурок из самого обычного листа бумаги, коим является японская техника оригами, занятие очень интересное и достаточно увлекательное. Однако сегодня, все больше и больше вызывает интерес модульное или, как его еще называют, объемное оригами, при котором фигурки складываются из многочисленных мелких деталей, так называемых модулей.Они могут иметь самую разнообразную форму. Модули удерживаются между собой за счет силы трения, что позволяет изгибать фигуру. Такое 3D-оригами стало широко распространенным не в одном только Китае, но и в других западных странах.
 Прижилось оно и у нас. Фигурки, сделанные при помощи модульного оригами будут не только оригинальным подарком Вашим близким, но и бесспорно украсят Ваш интерьер. В Японии такие фигурки вообще считаются талисманами.
Прижилось оно и у нас. Фигурки, сделанные при помощи модульного оригами будут не только оригинальным подарком Вашим близким, но и бесспорно украсят Ваш интерьер. В Японии такие фигурки вообще считаются талисманами.В данной технике каждая часть являет собой отдельный модуль, который делается из одного обычного листа бумаги. В большинстве своем это треугольные модули из которых можно сделать практически все, что Вам угодно. Делают модули из прямоугольных кусочков бумаги размером 53х74 мм или 37х53 мм. Затем, заранее подготовленные бумажные прямоугольники, нужно сложить вдвое, вдоль их длинных сторон и пополам, вдоль их коротких сторон.
Далее разворачиваем прямоугольники вторым сгибом к себе. Следуя линиям, сворачиваем края к центру, переворачиваем и загибаем нижние отвороты наверх.
Разгибаем, сложив треугольники по намеченным ранее линиям, и снова загибаем отвороты наверх. Складываем пополам и модуль готов. Затем один модуль всовывается в другой, при этом мы получаем целое произведение искусства.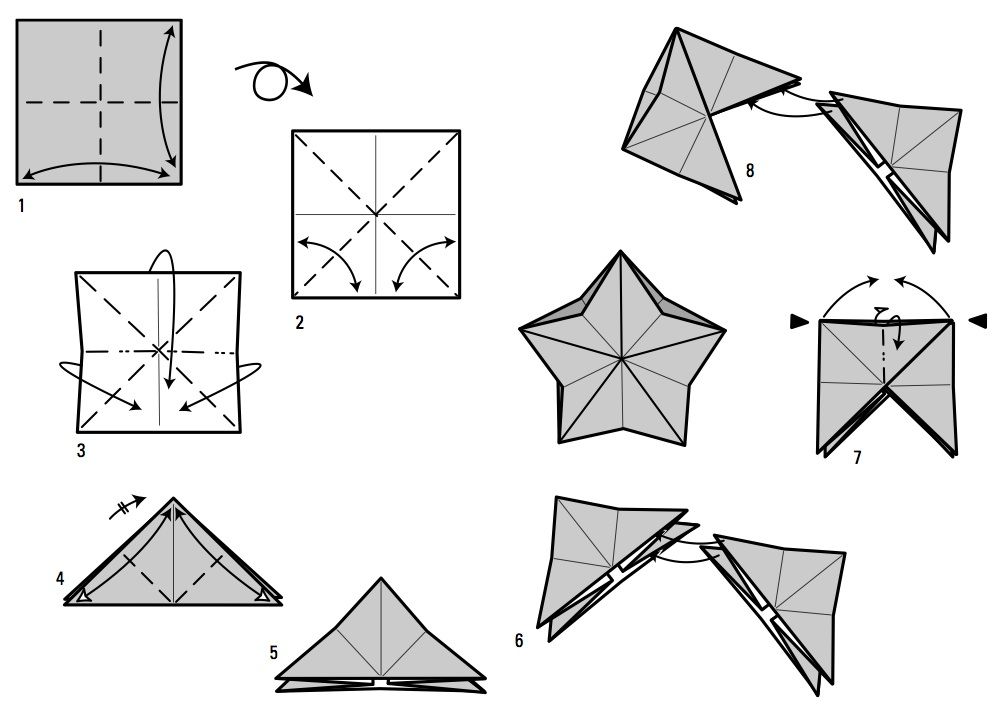 Зачастую модульные оригами схемы предполагают использование цветной бумаги, что придает готовому изделию особую оригинальность и элегантность.
Зачастую модульные оригами схемы предполагают использование цветной бумаги, что придает готовому изделию особую оригинальность и элегантность.
Одной из самых легких и широко используемых поделок является фигурка модульного оригами лебедь. При сравнительной простоте своей сборке, она имеет необычайно красивый внешний вид.
Также вы можете изготавливать различные цветы, вазы, героев любимых мультфильмов и много другое. Однако следует помнить, что если поделка отличается своей масштабностью, без использования клея Вам не обойтись, так как состоять она может из сотен, а то и тысяч модулей. В любом случае, соединяя модули, мы можем получить разнообразные конструкции. При этом фигура может быть как плоской, так и трехмерной.
Такое занятие считается развивающим и полезным для детей, ведь оно требует повышенного внимания и одновременно приводит в действие оба полушария мозга. Наряду с этим, это кропотливая и достаточно филигранная работа, требующая максимальной усидчивости и аккуратности.
Модульное оригами лебедь из треугольных модулей
Это одна из наиболее простых схем лебедя из бумажных треугольников. Для создания такого лебедя нам понадобится 458 белых модулей и один красный (или оранжевый) для клюва. Как сделать треугольный модуль>>
Итак приступим. Начинаем делать первые два ряда. Берем три модуля и располагаем их таким образом
Вставляем уголки двух модулей в кармашки третьего
Берем еще два модуля и подсоединяем к получившейся конструкции
Потом, точно также подсоединяем еще два модуля.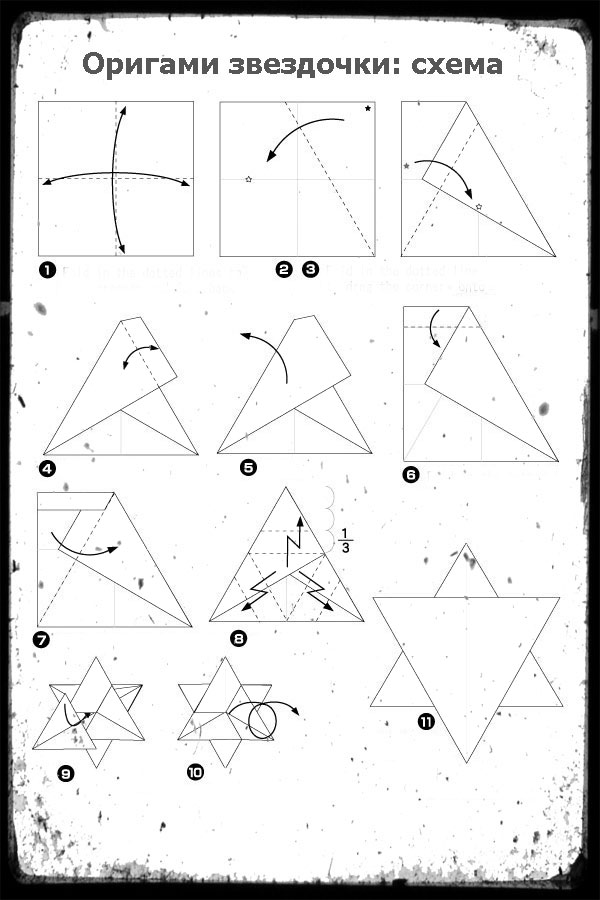 .. и понимаем что конструкция хилая и начинает разваливаться в руках при каждом неосторожном движении. Не расстраивайтесь, для того, чтобы все это держалось крепче, мы будем сразу собирать три ряда
.. и понимаем что конструкция хилая и начинает разваливаться в руках при каждом неосторожном движении. Не расстраивайтесь, для того, чтобы все это держалось крепче, мы будем сразу собирать три ряда
Для этого берем модуль и располагаем его вот так
И повторяем трюк вставки уголков в кармашки
Таким макаром собираем три ряда. Каждый ряд состоит из 30 модулей. И замыкаем их в круг.
Дальше, аналогично третьему ряду надеваем модули четвертого и пятого ряда
Теперь берем двумя руками слева и справа полученную юбочку, большими пальцами аккуратно вдавливаем середину, выворачивая изделие
Вместо юбочки получаем блюдечко
Края аккуратно загибаем вверх придавая изделию вот такую форму:
Вид сбоку
Вид снизу
Дальше надеваем шестой ряд модулей таким же образом как и предыдущий (не забывая что модули располагаются в шахматном порядке)
С седьмого ряда начинаем делать крылья. Таким же образом как и вредыдущих рядах одеваем 12 модулей. Потом пропускаем два уголка (от двух соседних модулей) и одеваем еще 12 модулей. На пропущенном месте с двумя уголками будет шея, а на более широком участке — хвост
Таким же образом как и вредыдущих рядах одеваем 12 модулей. Потом пропускаем два уголка (от двух соседних модулей) и одеваем еще 12 модулей. На пропущенном месте с двумя уголками будет шея, а на более широком участке — хвост
В следующем ряду уменьшаем каждое крыло на один модуль. Сответственно в каждом крыле восьмого ряда будет по 11 модулей
Вид сбоку
Так уменьшаем каждый ряд крыльев на один модуль, пока он всего один не останется (по ходу выкладывания крыльев слегка их выгибаем, придавая нужную форму)
поворачиваем лебедя к себе задом и делаем хвостик (тоже по принципу уменьшения на один модуль в каждом ряду)
Осталось сделать шею и голову лебедя. Для нее нам понадобится 19 белых и один красный модуль. В красном лучше заранее склеить уголки, чтобы клюв у лебедя не был раздвоенным. Собирается шея несколько по другому. Берем два модуля и располагаем вот так
Вставляем уголки одного в кармашки второго
и точно так же собирем все остальные, выгибая при сборке нужным образом
Теперь аккуратно насаживаем шею на оставленные два углка. Лебедь готов.
Лебедь готов.
Автор МК — Елена Соколова.
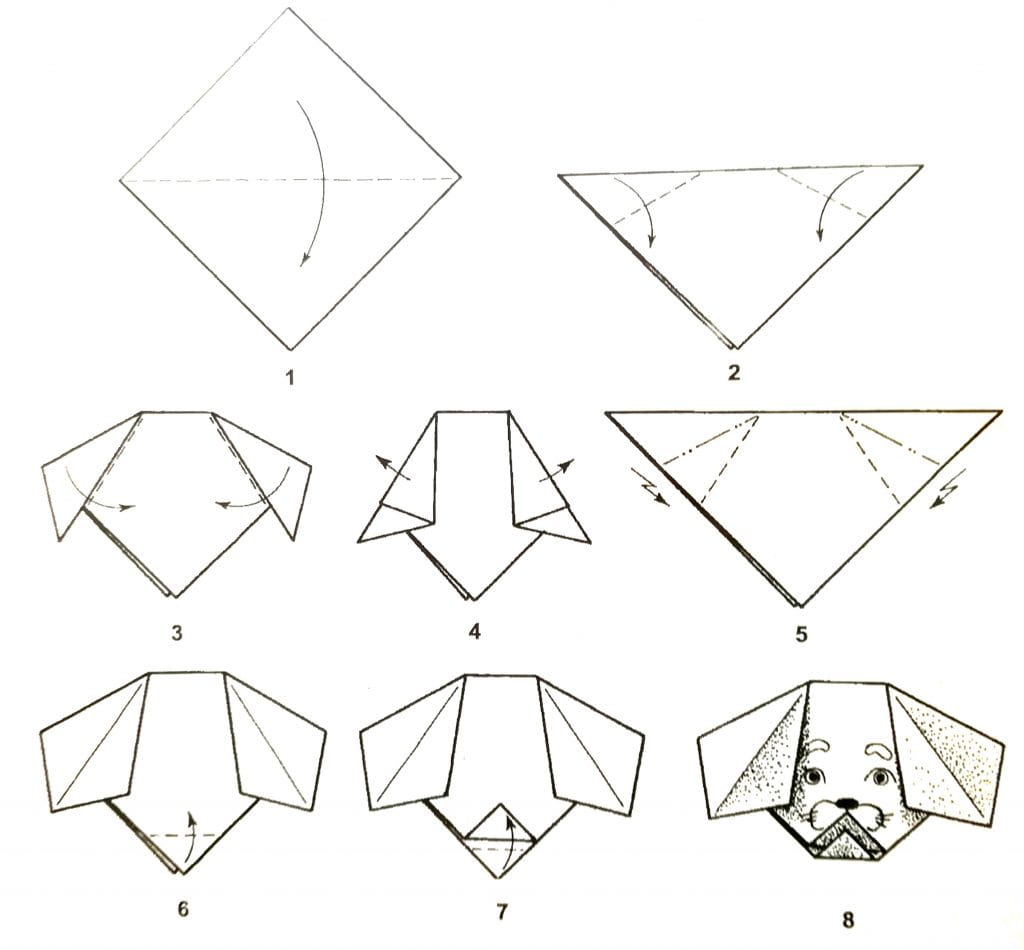
фигуры из бумаги для начинающих. Как сделать треугольник и квадрат по схеме? Сложные и легкие поделки
Техника создания фигурок оригами является безумно популярной уже много лет. Из бумаги можно создать множество красивых и оригинальных фигурок, не пользуясь клеевыми составами. Особенно эффектными получаются объемные оригами-поделки.
Что необходимо?
Техника моделирования оригами, позволяющая создавать фигурки самых разных видов, пользуется очень большой популярностью. Дело в том, что для создания любых самоделок мастеру не нужно подготавливать большое количество компонентов и инструментов. Самый главный элемент, который нужно будет использовать, – это бумага.
Самый главный элемент, который нужно будет использовать, – это бумага.
Существует несколько разновидностей бумаги в виде квадратов или прямоугольников, которые могут применяться для формирования красивых объемных фигурок-оригами.
- Белая бумага для принтера. Подобный материал является лучшим решением для реализации техники оригами. Эта бумага характеризуется высоким уровнем плотности, имеет шероховатую поверхность, прекрасно удерживает форму.
- Цветная бумага для принтера. Подобный материал не подвержен истиранию на сгибах. Эта бумага очень широко применяется при моделировании различных фигурок в технике модульного оригами.
- Стикеры. Такое название принадлежит небольшим по размерам разноцветным листикам, которые тоже очень хорошо подходят для создания разных поделок.
- Цветная бумага для творчества. Если запланировано создание качественной объемной фигурки-оригами, то подобный материал использовать не следует.
 Проблема в том, что этот вид бумаги является слишком тонким и мягким, на сгибах приобретает белый цвет.
Проблема в том, что этот вид бумаги является слишком тонким и мягким, на сгибах приобретает белый цвет. - Фольгированные бумажные листы. Отличаются высокой плотностью, прочностью. Из них можно мастерить сложные и замысловатые композиции. Материал подходит для скручивания.
- Листы из журналов. Неплохой вариант для изготовления фигурки в технике оригами. Подобный вид бумаги является достаточно плотным, легко поддается сгибам, хорошо держит форму.
- Денежные купюры. Их тоже можно использовать, но для создания маленьких поделок. Часто подобные изделия преподносят в качестве оригинального подарка.
- Специальная бумага для оригами. Реализуется в виде наборов. Бывает разноцветной, имеющей различные рисунки. Встречаются и экземпляры с разной текстурой.
- Калька. Может подойти для моделирования максимально простых вариантов для начинающих мастеров. Рекомендуется использовать кальку в качестве пробного материала.
 Проблема в том, что этот вид бумаги является слишком тонким и мягким, на сгибах приобретает белый цвет.
Проблема в том, что этот вид бумаги является слишком тонким и мягким, на сгибах приобретает белый цвет. Рекомендуется использовать кальку в качестве пробного материала.
Рекомендуется использовать кальку в качестве пробного материала.Подобрав подходящий материал для моделирования поделки, следует подготовить рабочее место. Все нужные составляющие желательно расположить так, чтобы они находились под рукой, а все ненужные детали лучше сразу отложить в сторону, чтобы они не мешали.
Общие принципы складывания
Прежде чем приступать к самостоятельному созданию оригинальной объемной фигурки-оригами, целесообразно узнать общие принципы складывания выбранной бумаги на примере модульных вариантов.
- Абсолютно любой тип конструкции объемного вида начинается с создания главного модуля. Классические варианты делаются треугольными. Чтобы придать материалу такую форму, его сгибают пополам по более длинной стороне, а верхние уголки подворачивают по направлению к центральной точке.
- Все имеющиеся выступы загибают сначала назад, а потом наверх. Уголочки требуется загибать во внутреннюю часть.
- Деталь сгибают как по вертикали, так и по горизонтали. Делать это следует так, чтобы кармашек был расположен на наружной стороне. В итоге должна получиться треугольная заготовка.
- Когда модули для формирования фигурки объемного типа будут готовы, их потребуется правильно соединить друг с другом. Самые простые разновидности оригами собирают буквально из одного листа.
- Схема сборки красивых конструкций является очень простой. Достаточно каждый модуль вставить в кармашек элемента, расположенного по соседству. Все элементы надежно удерживаются посредством трения последующих фиксирующихся деталей.
- Классический способ создания объемных фигурок-оригами предусматривает крепление модулей посредством насаживания на заостренные уголочки двух предыдущих заготовок на кармашек.
 Чтобы придать материалу такую форму, его сгибают пополам по более длинной стороне, а верхние уголки подворачивают по направлению к центральной точке.
Чтобы придать материалу такую форму, его сгибают пополам по более длинной стороне, а верхние уголки подворачивают по направлению к центральной точке.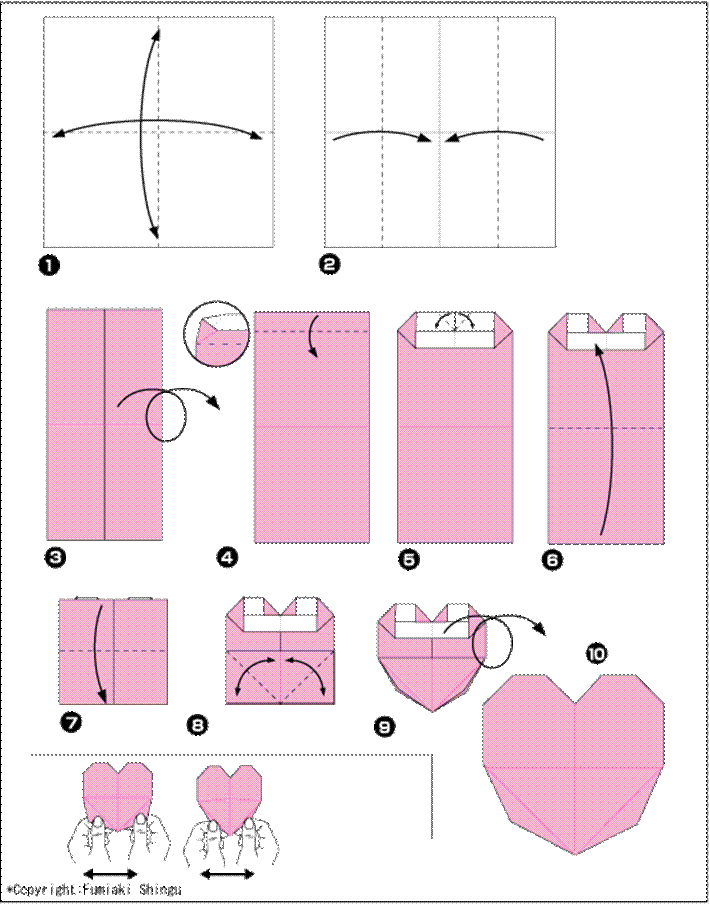
Как делать разные фигуры?
Объемная техника оригами позволяет создавать фигурки различных вариаций. Существует огромное количество интересных схем, с помощью которых своими руками удается моделировать из бумаги разнообразные цифры, буквы, животных, птиц, замки, домики и так далее.
Простые
Для начинающих мастеров, которые пока еще мало знакомы с техникой оригами, существуют максимально простые и понятные схемы моделирования объемных самоделок. К примеру, это могут быть фигурки в форме яблока, тыквы, звездочки.
Рассмотрим инструкцию по самостоятельному изготовлению красивой и простой объемной фигурки зеленого яблока. Для проведения работ необходимо подготовить бумагу двух цветов – светло-зеленого и травяного. Из первого материала надо вырезать квадрат, одна сторона которого составит 21 см. Из второго бумажного листа также понадобится вырезать квадратик 7х7 см.
- Сперва нужно сложить форму-базу будущей поделки. Это должен быть пятиугольник, разделенный на 10 одинаковых секций. Для этого можно воспользоваться подробной схемой.
- Полученный пятиугольник надо будет раскрыть. Все 5 диагоналей, что соединяют вершинки фигуры, понадобится сложить горой. Необходимо распределить 2 секции вправо и 3 налево. Это нужно, чтобы получить равнобедренный треугольник.
- Заготовку кладут перед собой, подгибают уголок справа, разравнивая его вдоль оси по центру.
- Далее раскрывают следующий треугольник. Его нужно сложить таким же образом. Необходимо двигаться по часовой стрелке, пока не будут загнуты все углы снизу на всех 5-и секциях.
- Складки потребуется тщательно разгладить. От этого будет зависеть надежность конструкции в дальнейшем.
- Нужно взять последнюю секцию. Ее поворачивают сложенной половинкой вниз, а потом подгибают вершинку.
- В отношении оставшихся 4-х секций понадобится провести такие же манипуляции. Затем необходимо изготовить крепеж, благодаря которому бумажное яблочко не будет разваливаться после наполнения воздухом.
- Следует открыть треугольник маленького размера и загнуть его свободную верхушку.
- Далее заготовку надо сложить в прежнем порядке. Треугольник сбоку потребуется раскрыть по типу кармашка и заложить туда незанятую верхушку (как при соединении шип-паз).
- Таким способом фиксируют все имеющиеся секции. На одинаковом расстоянии друг от друга потребуется их распределение.
- Заготовку необходимо поднести ко рту и дунуть в имеющееся отверстие. После этого поделка приобретет нужный объем. Нужно проверить качество заправки всех клапанов. Подправляют впадинки, если они есть.
- Теперь нужно сделать хвостик с зеленым листочком. Для этого берут темно-зеленый квадратик, сгибают по диагонали. Рисуют половинку листка и вырезают ее контур.
- Затем лист складывают повторно, косо подгибают верхушку. После маленького отступа формируют еще одну складку. Так нужно делать до заполнения всего листка.
- Готовую деталь вставляют в дырку, через которую надувалось яблочко. Из темно-зеленой бумаги можно вырезать длинную полоску треугольной формы и закрутить ее в жгутик. Этот элемент станет яблочным хвостиком. Лист и хвостик надо будет склеить.

Мастерится это изделие не так сложно, как может показаться на первый взгляд.
Сложные
Существует очень много более сложных схем изготовления объемных самоделок. Часто таким образом мастера делают разных животных, к примеру, кота, кенгуру, пингвина, кита и так далее.
Рассмотрим, как сделать сложную фигурку цыпленка посредством модульной техники оригами.
- Понадобится сделать 221 модуль желтого цвета, 343 белых модуля, 1 красный модуль для клюва птички.
- Сперва нужно сомкнуть круг из первых 16 модулей желтого цвета.
- Стартовый ряд следует нарастить 4 из 16 желтыми деталями.
- Потом собирают еще 6 рядов. Модули должны быть направлены гипотенузой наружу.
- Когда туловище цыпленка будет сделано, нужно смастерить крылья. Каждое из них делается из 6 желтых модулей. Сперва соединяют первые 3, а потом следующие 2 модуля и последний 1 модуль.
- Крылышки фиксируют на боках туловища, делая одинаковые расстояния. По центру следует вставить деталь-клюв.
- Собирают скорлупу. Для этого смыкают кольцо из 16 белых деталей. Поверх наращивают еще 4 ряда из 14 белых элементов. В 5 и 6 рядах добавляют по 7 модулей.
- Верхушка скорлупы делается из 3-х рядов в 8 модуле. 4-й ряд увеличивают еще на 16 деталей.
- Из бумаги можно сделать глазки цыпленка и приклеить их к готовому туловищу. После прикрепления скорлупы красивая поделка в модульной технике оригами будет готова.
О том, как своими руками сделать объемное сердечко оригами, смотрите в следующем видео.
6 лучших схем оригами, идеи и пошаговые инструкции (фото + видео)
Фото: blog.peacerevolution.netМилые и романтичные сердечки из бумаги – трогательный жест внимания, который порадует близких. Ими можно дополнить подарок, положить внутрь маленький сувенир или написать на них необычную записку. Рассказываем, как быстро и легко сделать аккуратную валентинку своими руками!
1. Сердечко оригами для начинающих
Возьми квадратный лист плотной двусторонней бумаги и визуально раздели его на три части вертикалями.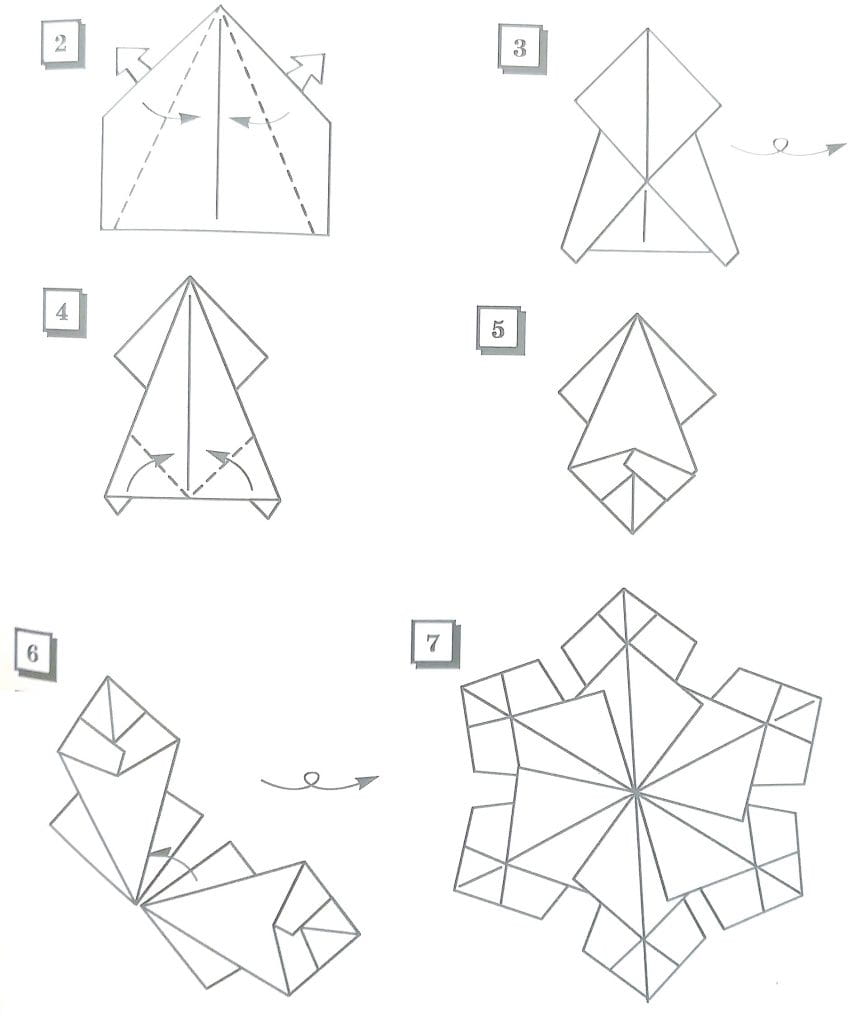 Заверни нижний правый угол на две трети, а потом четко по нему подогни обе стороны на треть. Разверни деталь углом-кармашком вниз и отогни вниз верхнюю половину нижнего слоя. Можешь расправить верхние половинки, чтобы они были более округлыми.
Заверни нижний правый угол на две трети, а потом четко по нему подогни обе стороны на треть. Разверни деталь углом-кармашком вниз и отогни вниз верхнюю половину нижнего слоя. Можешь расправить верхние половинки, чтобы они были более округлыми.
2. Сердце из бумаги А4
Разметь вертикаль и горизонталь цветного листа бумаги А4 и заверни верхние углы в «домик». Подогни нижние углы у основания, чтобы получился ромб, и переверни заготовки на обратную сторону. Сформируй половинки сердца со всех сторон и снова переверни деталь – готово!
Фото: idoska-yaroslavl.ruОригами из бумаги для начинающих: 10 легких схем
3. Объемное сердце из бумаги
Квадратный лист плотной бумаги сложи пополам, чтобы разметить его оси. Положи прямоугольник сгибом вниз и подверни его края вверх, немного не доходя до середины. Еще раз заверни углы, выровняй и подтяни их к центру, как на фото.
Каждый нижний сегмент разверни в вытянутый пятиугольник-лепесток – это будут боковые стороны сердца. Переверни заготовку и сформируй верхние округлости, вжимая к центру фигуры. Надуй сердечко через маленькое отверстие в остром носике.
Переверни заготовку и сформируй верхние округлости, вжимая к центру фигуры. Надуй сердечко через маленькое отверстие в остром носике.
4. Сердце-закладка для книг
Возьми прямоугольный лист яркой упаковочной бумаги и сложи его пополам однотонной стороной внутрь. Обе стороны отогни «крыльями» вверх посередине треугольника – у тебя получится своеобразный перевернутый домик. Переверни заготовку, а затем сформируй округлости сердца, загибая углы – и уже можешь надевать его прямо на страницу!
Фото: schoolmum.net Фото: paper-land.ruКак сделать ракету из бумаги: 10 пошаговых схем
5. Бумажное сердечко с крыльями
Тебе понадобится квадратный лист бумаги с яркой лицевой стороной и белой изнанкой. Разметь его оси, подверни к середине нижний край и разверни деталь на 180 градусов. Подогни треугольник по бокам, чтобы сделать трапеции, и снова переверни деталь белой стороной кверху.
Сложи «домик», как на схеме, разверни заготовку цветной вершиной вниз и загни верхний слой вверх. Выпрями его в длинную вытянутую трапецию и сформируй из нее крылышки обычной лесенкой. Хорошо пережми их по центру, чтобы придать форму, и разверни сердце лицом обратно.
Фото: propovyazki.ru6. Оригами сердце с цветком
Это оригами сердечко из бумаги кажется очень сложным, но достаточно начать его складывать, чтобы понять, что не все так страшно. Только заранее запасись тонкой плоской палочкой от эскимо или пилочкой для ногтей, чтобы распрямлять лепестки.
Отрежь бумажный прямоугольник с соотношением сторон 2:1, разметь все середины, четверти и диагонали, как на схеме. Левую половину квадрата сложи по треугольникам, а углы его верхнего слоя разверни в квадраты. Загни бока квадрата к середине и каждый из них разложи в ромб-лепесток с помощью тонкого инструмента.
Второй такой же лепесток разверни снизу, а потом повтори все по кругу для другой половины сердца. При желании спрячь внутрь боковые углы фигуры, чтобы сделать ее более округлой. Изящное объемное сердце с цветком посередине готово!
При желании спрячь внутрь боковые углы фигуры, чтобы сделать ее более округлой. Изящное объемное сердце с цветком посередине готово!
8 лучших схем, как сделать коробку из бумаги
Видео: Оригами сердце с посланием — пошагово
Понравилась публикация? Подпишись на наш канал в Яндекс.Дзен, это очень помогает нам в развитии!
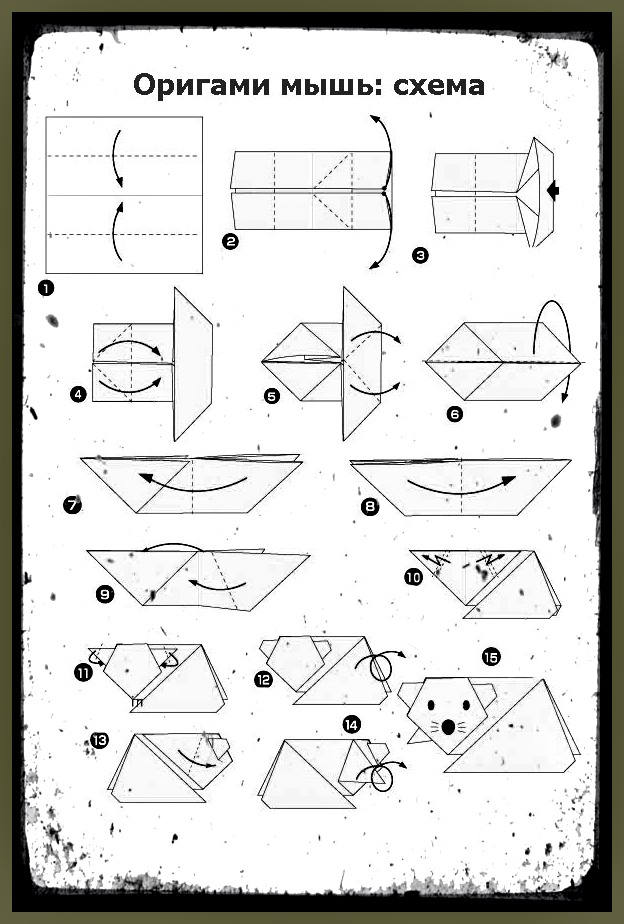
ОРИГАМИ ИЗ ТРЕУГОЛЬНЫХ МОДУЛЕЙ | Треугольное оригами
Модульное оригами представляет собой удивительную технику создания фигурок из бумаги с помощью нескольких одинаковых деталей или треугольных модулей. Освоив хотя бы некоторые приемы их соединения, можно создавать объемные модели. Это могут быть оригами из треугольных модулей -разнообразные животные, птицы, цветы,сказочные персонажи, дома… Очень часто встречаются схемы оригами, использующие треугольные модули.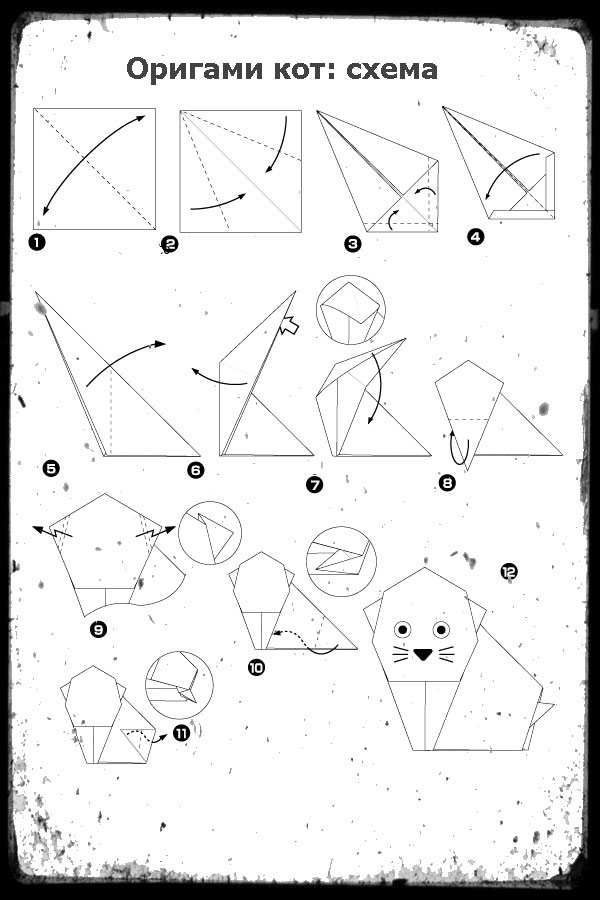 Как сделать треугольное оригами описано ниже.
Как сделать треугольное оригами описано ниже.
Для создания оригами из треугольных модулей можно выбирать практически любую бумагу – принтерную, газетную, журнальную и пр. Сложить треугольный модуль не трудно. Немного опыта – и сделать это можно с закрытыми глазами. Благодаря этим модулям можно создавать фигурки любых размеров и любой сложности.
Чтобы сделать треугольный модуль для оригами, нужно нарезать бумагу маленькими равными прямоугольниками. Их размер напрямую зависит от размера планируемой фигуры. Для удобства можно обычный лист А4 разрезать на 16 частей, разлиновав каждую его сторону на 4 равных отрезка. Аналогичным образом лист можно разделить на 32 части, если длинную сторону листа разлиновать на 8 равных отрезков.
При желании можно выбрать любой другой подходящий размер, но лучше все-таки пользоваться одним и тем же, чтобы в случае необходимости можно было разобрать фигуру и создать что-нибудь новое из тех же модулей. Для нарезки удобно использовать канцелярский нож или специальный резак для бумаги.
Чтобы сделать треугольный модуль для оригами, нужно:
1. согнуть бумажный прямоугольник пополам вдоль;
2. согнуть пополам, чтобы наметить линию, и разогнуть обратно;
3. верхние уголки загнуть внутрь вдоль намеченного центрального сгиба;
4. перевернуть на обратную сторону;
5. загнуть нижнюю часть детали вверх;
6. загнуть «торчащие» уголки за деталь, чтобы образовался треугольник;
7. отогнуть обратно нижнюю часть;
8. загнуть нижнюю часть опять вверх, чтобы «торчащие» уголки оказались внутри;
9. сложить треугольник пополам наподобие книжки.
Сделав все правильно, получается треугольник с двумя нижними кармашками. Впоследствии фигурки образуются путем вкладывания угла одного модуля в кармашек другого.
В схемах оригами из треугольных модулей используются интуитивно понятные условные обозначения, делающие их универсальными и доступными для людей независимо от их национальной принадлежности и владения иностранными языками.
Существуют две принципиальные схемы, называемые «согнуть долиной» и «согнуть горой». Различаются они тем, где находится образовавшаяся линия сгиба, не доходящая до края фигурки. «Согнуть долиной» — значит, меньшая часть располагается ближе к мастеру. «Согнуть горой» — меньшая часть располагается дальше, как бы скрываясь за большей частью.
Делаем модули треугольной формы,с помощью которых создаются изящные поделки из бумаги
Оригами из треугольных модулей — Мастер класс
Следующая статья :
Простые оригами поделки для детей
вернуться на Главную страницу
Поделки из бумаги в технике оригами. Фото галерея
ТАКЖЕ УЗНАЙТЕ…
Следующие материалы:
Предыдущие материалы:
Модульное оригами для начинающих или с чего начать?
Один мудрец однажды сказал что…
«…Красота – дело рук Бога и человека.
 Бог дает вдохновение и образы. А человек, движимый вдохновением, черпает образы из окружающей красоты всего сущего – и создает шедевр».
Бог дает вдохновение и образы. А человек, движимый вдохновением, черпает образы из окружающей красоты всего сущего – и создает шедевр».
Модульное оригами – это красивое хобби. Это поделки, созданные руками человека, на основе тех образов, которые он хотел бы воплотить в реальность.
Сегодня все больше и больше людей влюбляются в это тонкое и такое простое искусство. Искусство, которому может научиться каждый — стоит только узнать, как сложить из бумаги модуль и как начинающим в модульном оригами собрать все в единую конструкцию.
Если вы до сих пор считаете, что для создания этих бумажных объемных фигур необходимы недюжие способности и глыба таланта – то вы преувеличиваете.
Модульное оригами даже дети 7-летнего возраста осваивают за 2 вечера (первый вечер они сворачивают из бумаги модуль, второй вечер учатся крепить их в единое целое).
Сегодня я расскажу Вам, начинающим в модульном оригами, как начать заниматься модульным оригами, как подобрать бумагу и где ее можно найти, как нарезать бумагу и главное, чем нарезать.
Посмотрите пожалуйста видеоролик «Возможности модульного оригами», у вас есть все шансы сделать, причем самостоятельно, такую красоту, а я вам в этом помогу.
Принципы сборки модульного оригами
Чтобы вам сразу стало понятно по каким законам существует модульное оригами, я хочу показать Вам, КАК ЭТО ПРОСТО – собирать объемные вещи из такого бумажного конструктора.
Вот смотрите: из бумажных модулей можно собрать обычные плоские фигуры, например, вот такую рамку для фотографии в форме сердечка. Кстати, если Вы посмотрите на рисунок выше (с лебедями), то увидите, что лебединые шеи делаются точно по такому же плоскостному принципу соединения модулей.
Для создания объемных фигур те же самые бумажные модули собираются уже по другим правилам соединения.
На самом деле сделать из модульного оригами ту или иную модель ДОСТАТОЧНО ЛЕГКО. При условии, что сделанные Вами бумажные модули изготовлены из правильной бумаги. Вот об этом, о бумаге, мы сейчас поговорим…
Какая бумага подходит для модульного оригами?
Но тут все очень просто: сейчас, в наше прогрессивное время, без бумажки не обойтись. Всех завалили бумажной волокитой и меня в том числе. Это я о том, что бумагу для начинающих в модульном оригами, можно купить в любом магазине канцелярии, в компьютерном магазине, да вообще везде она есть.
Для начала нужна белая бумага, лучше всего подходят марки «Снегурочка» и «SvetoCopy», продаются они пачками по 500 листов, есть пачки и меньшего размера. Они не дорогие, в пределах от 150р. — 200р. за пачку (в зависимости от региона страны).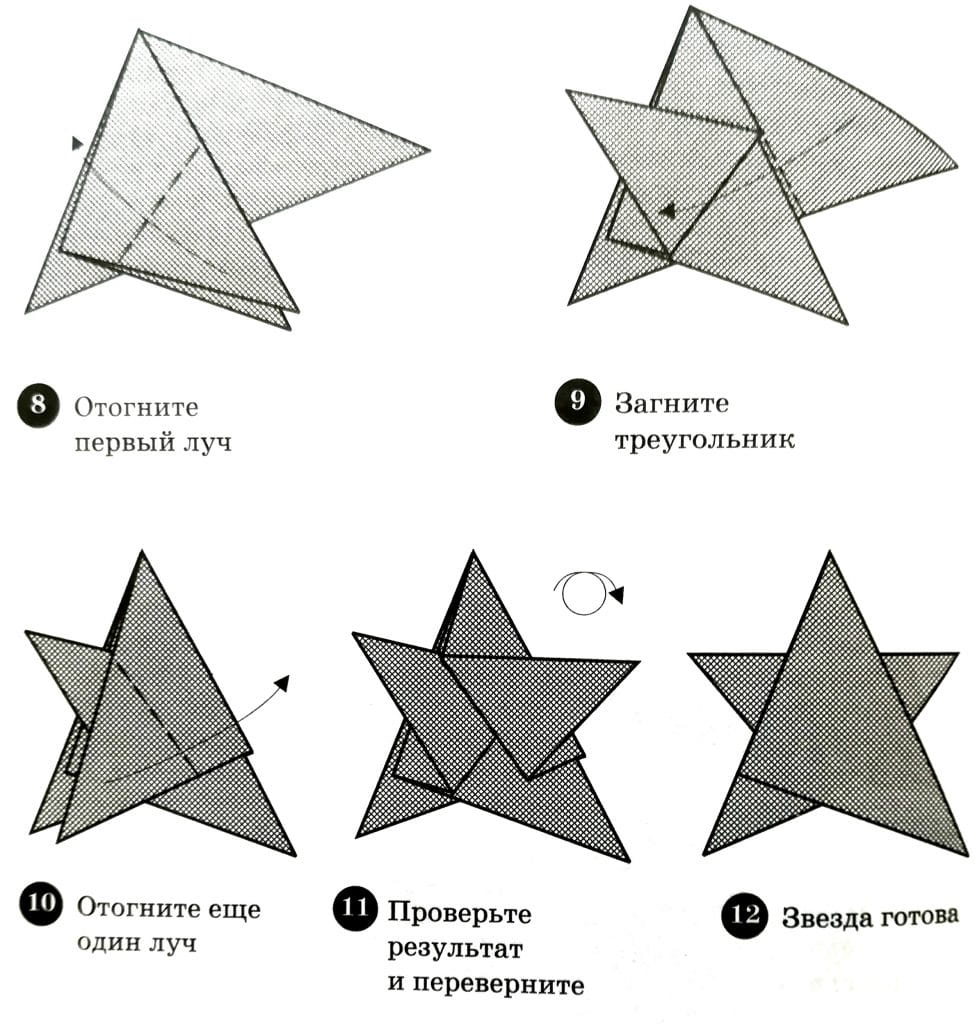 Плотность должна быть 80 г/м2, ее Вы можете найти на упаковке. Размер бумаги стандартного размера — А4 (в некоторых случаях есть бумага размером А3).
Плотность должна быть 80 г/м2, ее Вы можете найти на упаковке. Размер бумаги стандартного размера — А4 (в некоторых случаях есть бумага размером А3).
А вот с цветной бумагой немного посложней, в том смысле, что она почти в три раза дороже. Марки цветной бумаги очень разнообразны, например: «Master Color», «Maestro Color», «Колорит», «Xerox», «IQ Color» и нашего производства – «Гознак», «Нота», «Радуга» и еще куча всяких марок. Они почти ничем не отличаются по качеству, по плотности и по яркости цветов.
Продается она пачками по 500 листов, есть пачки поменьше, размером А4 или в некоторых случаях А3. Для удобства покупателям многие продавцы магазинов продают цветную бумагу поштучно или делают наборы из нескольких цветов. Плотность 80 г/м2, а вот цена может колебаться от 550р. до 700р. за пачку, может и выше (в зависимости от региона страны). Есть еще много вариантов цветной бумаги — это может быть бумага для стикеров, для аппликации, для поделок и т. д., но об этом мы поговорим позже. Самый лучший вариант и оптимальный — это те виды бумаги которые я перечислил выше.
д., но об этом мы поговорим позже. Самый лучший вариант и оптимальный — это те виды бумаги которые я перечислил выше.
Классификация цветной бумаги [КОДЫ]
Для того, чтобы Вам было удобнее выбирать цвет бумаги или заказывать по интернету или в магазине, предлагаю Вашему вниманию схемы КОДОВ.
К каждому цвету соответствует свой код — это очень удобно, для заказа бумаги.
Я частенько пользуюсь этими схемами, они выручают в различных ситуациях
В следующей статье Вы узнаете, как лучше нарезать бумагу, чем и какие еще инструменты и приспособления нужны начинающему в модульном оригами.
Origami привлекла внимание сообщества микропроизводителей, поскольку оно позволило использовать такие приложения, как микрозеркала, устройства для инкапсуляции и микрозахваты. Тем не менее, методы изготовления систем оригами, доступные в миллиметровом и субмиллиметровом масштабе, имеют узкую область применения со стратегиями синтеза для конкретных продуктов.Поэтому обобщенные методы изготовления оригами в таких масштабах, имитирующие аддитивное производство, представляют большой интерес. Небольшие масштабы длины этих систем требуют реализации методов самоскладывания или складывания с помощью по мере приближения к изготовлению. В этой работе метод самостоятельного складывания и вспомогательный метод складывания демонстрируются как стратегии изготовления оригами. Однослойные пленки фотополимера с узорчатыми свойствами вдоль их плоскостной области используются для создания плоских предшественников структур оригами.Свойства пленок моделируются путем контроля плотности их сшивания с помощью процесса фотолитографии с двойной экспозицией. Для вспомогательного складывания в качестве исполнительного механизма используется поверхностное натяжение капли. Податливые области складок, изготовленные из менее сшитого фотополимера, соединяют относительно более жесткие области лица. Тем не менее, методы изготовления систем оригами, доступные в миллиметровом и субмиллиметровом масштабе, имеют узкую область применения со стратегиями синтеза для конкретных продуктов.Поэтому обобщенные методы изготовления оригами в таких масштабах, имитирующие аддитивное производство, представляют большой интерес. Небольшие масштабы длины этих систем требуют реализации методов самоскладывания или складывания с помощью по мере приближения к изготовлению. В этой работе метод самостоятельного складывания и вспомогательный метод складывания демонстрируются как стратегии изготовления оригами. Однослойные пленки фотополимера с узорчатыми свойствами вдоль их плоскостной области используются для создания плоских предшественников структур оригами.Свойства пленок моделируются путем контроля плотности их сшивания с помощью процесса фотолитографии с двойной экспозицией. Для вспомогательного складывания в качестве исполнительного механизма используется поверхностное натяжение капли. Податливые области складок, изготовленные из менее сшитого фотополимера, соединяют относительно более жесткие области лица. При нанесении капли на пленки включается изгибная деформация, локализованная в податливых складках. Для метода самоскладывания процесс фотолитографии разработан таким образом, что раствор проявителя неоднородно поглощается по всему поперечному сечению областей сгиба.Эта неоднородность концентрации приводит к локальному изгибу, когда проявитель высвобождается при нагревании. Этот метод самоскладывания на основе диффузии позволяет программировать сгибы для достижения заданных углов сгиба. Оба метода, обеспечиваемые узорчатыми пленками, изготовленными с использованием фотолитографии с двойной экспозицией, обещают возможность изготовления миллиметровых и субмиллиметровых систем оригами произвольной формы. При нанесении капли на пленки включается изгибная деформация, локализованная в податливых складках. Для метода самоскладывания процесс фотолитографии разработан таким образом, что раствор проявителя неоднородно поглощается по всему поперечному сечению областей сгиба.Эта неоднородность концентрации приводит к локальному изгибу, когда проявитель высвобождается при нагревании. Этот метод самоскладывания на основе диффузии позволяет программировать сгибы для достижения заданных углов сгиба. Оба метода, обеспечиваемые узорчатыми пленками, изготовленными с использованием фотолитографии с двойной экспозицией, обещают возможность изготовления миллиметровых и субмиллиметровых систем оригами произвольной формы. |
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка браузера на прием файлов cookie
Существует множество причин, по которым файл cookie не может быть установлен правильно. Ниже приведены наиболее распространенные причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки браузера, чтобы принять файлы cookie, или спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файл cookie.
- Ваш браузер не поддерживает файлы cookie. Попробуйте другой браузер, если вы подозреваете это.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы это исправить, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
 Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Предоставить доступ без файлов cookie потребует от сайта создания нового сеанса для каждой посещаемой вами страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в файле cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только та информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт
не может определить ваше имя электронной почты, если вы не решите ввести его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступ к
остальной части вашего компьютера, и только сайт, создавший файл cookie, может его прочитать.
Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступ к
остальной части вашего компьютера, и только сайт, создавший файл cookie, может его прочитать.
Аддитивный алгоритм для дизайна оригами
Значение
Оригами, искусство складывания бумаги, является новой платформой для механических метаматериалов.Предыдущая работа по проектированию структур на основе оригами была сосредоточена на простых геометрических конструкциях для ограниченных пространств типологий оригами или глобальных задачах оптимизации с ограничениями, которые трудно решить. Здесь мы меняем математическую, вычислительную и физическую парадигму дизайна оригами, предлагая простой подход к локальному маршированию, который приводит к конструктивной теореме о геометрии совместимых направлений роста на границе данного сложенного семени. Мы показываем, как это открытие приводит к простому алгоритму аддитивного проектирования всех развертываемых четырехугольных поверхностей, что позволяет проектировать новые складные метаструктуры из плоских листов в любом масштабе.
Ключевые слова: оригами, вычислительный дизайн, метаматериалы, аддитивное производство
Abstract
Вдохновленные очарованием аддитивного производства, мы ставим проблему дизайна оригами с другой точки зрения: как мы можем вырастить складчатую поверхность в трех измерениях из семени так, чтобы оно гарантированно было изометрично плоскости? Мы решаем эту задачу в два этапа: сначала выявляя геометрические условия согласованного завершения двух отдельных складок в единую развертывающуюся четверную вершину, а затем показывая, как это основание позволяет вырастить геометрически согласованный фронт на границе заданной складчатости. семя.Это дает полный маршевый или аддитивный алгоритм для обратного проектирования полного пространства развертываемых четырехъядерных шаблонов оригами, которые можно сложить из плоских листов. Мы иллюстрируем гибкость нашего подхода, выращивая упорядоченные, неупорядоченные, прямые и криволинейно-сложенные оригами и подгоняя поверхности заданной кривизны с помощью сложенных аппроксимаций.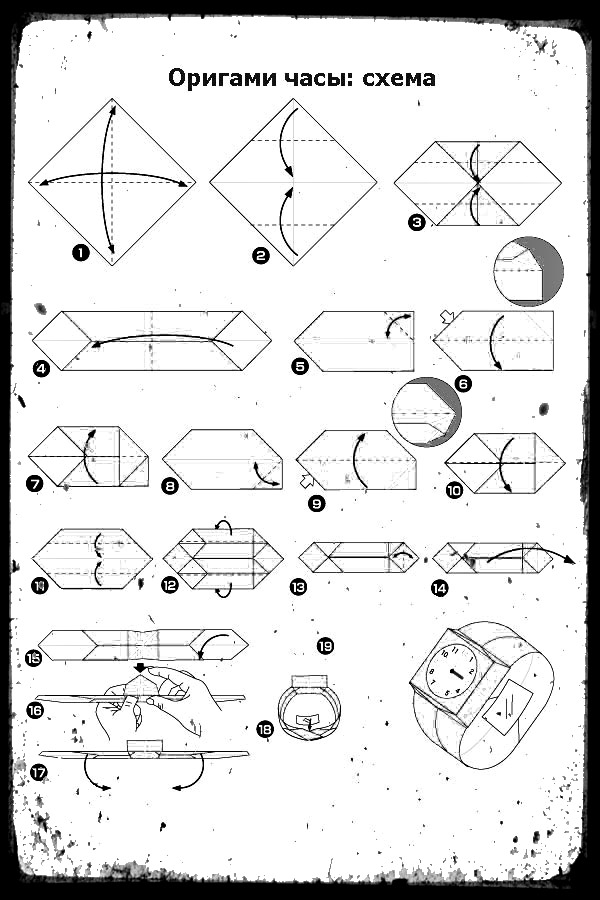 В целом, наш простой переход от глобального поиска к локальному правилу может изменить дизайн метаструктуры на основе оригами.
В целом, наш простой переход от глобального поиска к локальному правилу может изменить дизайн метаструктуры на основе оригами.
Складные узоры возникают в природе в виде систем, включающих крылья, листья и кишки насекомых (1–4), и имеют долгую историю в декоративных, церемониальных и педагогических традициях оригами по всему миру.Совсем недавно они начали привлекать внимание математиков, увлеченных моделями и пределами складчатости (5–9), а также инженеров и ученых, вдохновленных их технологическими перспективами (10–18).
Простейшее оригами представляет собой одну вершину с четырьмя складками, своеобразный атом водорода складок ровно с одной внутренней степенью свободы (DOF). Узоры, состоящие из четырехкоординатных вершин и четырехугольных граней, называются четырехугольными оригами, которые могут иметь изолированные складчатые конфигурации, изометричные плоскости, если их вообще можно сложить.Механическая реакция конструкций и материалов, полученных из четырехугольного оригами, в значительной степени определяется геометрическими нарушениями, возникающими при складывании. Использование этих шаблонов для программирования жестко-складываемых и плоских складных, гибких или мультистабильных систем требует учета дополнительных симметрий (19) и складок (20), что делает четырехъядерное оригами многообещающей платформой для метаструктур любого масштаба, от наноскопического до архитектурного. . Это вызвало значительный научный интерес к проблеме их дизайна, но задача поиска четырехугольных узоров, которые действительно складываются, или, наоборот, разворачивающихся поверхностей, имеет ограниченные решения произвольной формы.
Использование этих шаблонов для программирования жестко-складываемых и плоских складных, гибких или мультистабильных систем требует учета дополнительных симметрий (19) и складок (20), что делает четырехъядерное оригами многообещающей платформой для метаструктур любого масштаба, от наноскопического до архитектурного. . Это вызвало значительный научный интерес к проблеме их дизайна, но задача поиска четырехугольных узоров, которые действительно складываются, или, наоборот, разворачивающихся поверхностей, имеет ограниченные решения произвольной формы.
Предыдущие исследования дизайна четырехъядерных оригами, как правило, были сосредоточены на мозаиках с периодической геометрией и конкретными назначениями гор / долин (MV), принятыми априори, с хорошо известным шаблоном Миура-ори (21), являющимся каноническим примером, и обычно использовали либо прямые геометрические методы для параметризации простых вариантов конструкции (22–30), либо алгоритмы оптимизации для обобщения известных типологий складывания (31–34). Первые обычно обеспечивают всестороннее понимание ограниченного пространства проектов, имеющих сильное качественное сходство, т.е.т. е. те, которые демонстрируют определенную симметрию, и включают конструкции, которые неизбежно зависят от конкретного случая. Последние обычно требуют кодирования нелинейных ограничений возможности разработки в невыпуклой, многомерной структуре оптимизации и используют хорошо известную периодическую схему складывания в качестве эмпирической отправной точки. Эти вычислительные методы являются общими, но у них есть две проблемы: сложность поиска хорошего предположения, обеспечивающего сходимость к желаемому локальному решению, и отсутствие масштабируемости для больших задач.Таким образом, в то время как многие современные стратегии использовались для объяснения широкого разнообразия моделей четырехъядерного оригами, общая проблема дизайна четырехъядерного оригами допускала только частичные решения, и наука по большей части следовала форме искусства.
Первые обычно обеспечивают всестороннее понимание ограниченного пространства проектов, имеющих сильное качественное сходство, т.е.т. е. те, которые демонстрируют определенную симметрию, и включают конструкции, которые неизбежно зависят от конкретного случая. Последние обычно требуют кодирования нелинейных ограничений возможности разработки в невыпуклой, многомерной структуре оптимизации и используют хорошо известную периодическую схему складывания в качестве эмпирической отправной точки. Эти вычислительные методы являются общими, но у них есть две проблемы: сложность поиска хорошего предположения, обеспечивающего сходимость к желаемому локальному решению, и отсутствие масштабируемости для больших задач.Таким образом, в то время как многие современные стратегии использовались для объяснения широкого разнообразия моделей четырехъядерного оригами, общая проблема дизайна четырехъядерного оригами допускала только частичные решения, и наука по большей части следовала форме искусства.
Вдохновленный простой операцией экструзии краев из вычислительного дизайна и аддитивного производства, можно задать следующую обратную задачу: как мы можем расширить границу сложенной четырехугольной поверхности оригами наружу так, чтобы новая поверхность оставалась развертываемой, т. е.е., изометричных плоскости и, таким образом, поддающихся изготовлению из плоских листов? В случае с оригами это означает, что процесс складывания может трансформировать узор через промежуточные конфигурации во второй глобальный минимум энергии, сконструированную складчатую поверхность. Хотя это обычно подразумевает геометрическое нарушение на промежуточных этапах, недавно было разработано несколько маршевых алгоритмов для проектирования четырехъядерных оригами, которые, помимо развертывания в плоскости, могут развертываться жестко, то есть без геометрического нарушения.Это позволяет разворачивать конфигурации от плоского к сложенному и к плоскосложенному с одной степенью свободы, подклассу развертываемого четырехъядерного оригами, известного как жесткое и плоское складывание (35–37), например, метод «головоломки» для проектирования жесткого складывания. четырехъядерное оригами (36), комбинаторная стратегия, заимствованная из художественного модульного дизайна оригами (38), в котором геометрически совместимые сложенные элементы выбираются из заранее определенного набора дискретных модулей для увеличения границ сложенной объемной модели.
е.е., изометричных плоскости и, таким образом, поддающихся изготовлению из плоских листов? В случае с оригами это означает, что процесс складывания может трансформировать узор через промежуточные конфигурации во второй глобальный минимум энергии, сконструированную складчатую поверхность. Хотя это обычно подразумевает геометрическое нарушение на промежуточных этапах, недавно было разработано несколько маршевых алгоритмов для проектирования четырехъядерных оригами, которые, помимо развертывания в плоскости, могут развертываться жестко, то есть без геометрического нарушения.Это позволяет разворачивать конфигурации от плоского к сложенному и к плоскосложенному с одной степенью свободы, подклассу развертываемого четырехъядерного оригами, известного как жесткое и плоское складывание (35–37), например, метод «головоломки» для проектирования жесткого складывания. четырехъядерное оригами (36), комбинаторная стратегия, заимствованная из художественного модульного дизайна оригами (38), в котором геометрически совместимые сложенные элементы выбираются из заранее определенного набора дискретных модулей для увеличения границ сложенной объемной модели.
Здесь мы принципиально отклоняемся от этих предыдущих подходов, предоставляя унифицированную структуру, которая идентифицирует полное непрерывное семейство совместимых складчатых полос, которые могут быть выдавлены непосредственно из границы складчатой модели.Это закладывает основу для алгоритма, который позволяет нам исследовать все пространство развертываемых четырехъядерных оригами, не ограничиваясь только жесткими и/или плоскими складными конструкциями. Мы начнем с изучения гибкости углов и длин, связанной со слиянием двух пар сложенных граней на общей границе, что дает условия геометрической совместимости для проектирования четырехкоординатного одновершинного оригами. Затем мы применяем одновершинную конструкцию, чтобы определить пространство совместимых четырехполосных оригами на границе существующей складчатой модели.Важно установить, что новые направления внутренних краев и расчетные углы вдоль фронта роста образуют одномерное семейство, параметризованное выбором ориентации одной грани в пространстве вдоль фронта роста. Результатом является аддитивный геометрический алгоритм для эволюции складчатых фронтов вокруг заданного начального числа в складчатую поверхность, устанавливающий средства для описания всего пространства проектирования общих четырехъядерных поверхностей оригами. Этот конструктивный алгоритм (39) обеспечивается следующим:
Результатом является аддитивный геометрический алгоритм для эволюции складчатых фронтов вокруг заданного начального числа в складчатую поверхность, устанавливающий средства для описания всего пространства проектирования общих четырехъядерных поверхностей оригами. Этот конструктивный алгоритм (39) обеспечивается следующим:
Теорема. Пространство направлений новых внутренних ребер по всему фронту роста в четверном оригами есть одно — размерное — т.е. , однозначно определяемое одним углом .
Доказательство: Наше доказательство в основном состоит из трех частей: построение одной вершины, построение смежных вершин и рост полного роста, подробности приведены в SI Приложение , раздел S1.
1.
Одновершинная конструкция: Предположим, что нам дана вершина вдоль фронта роста с вектором положения xi (), с двумя соседними вершинами фронта роста, обозначенными xi−1,xi+1.
Обозначим два проектных угла границы, падающие на xi на существующей поверхности, через θi,3 и θi,4, а угол в пространстве в точке xi вдоль фронта роста обозначим через βi=∠{−ei,ei+1}∈(0 ,π) (), где ei=xi−xi−1 и ei+1=xi+1−xi. Чтобы получить новый вектор направления ребра ri, который задает направление внутреннего ребра [xi,xi′] на расширенной четырехугольной поверхности оригами, пусть αi∈[0,2π) будет левосторонним углом закрылка вокруг ei от плоскость βi в плоскость нового четырехугольника, содержащего ri и ei (). Заметим, что одновершинное оригами в точке xi удовлетворяет условию развертываемости суммы локальных угловПостроение четверного оригами.( A ) Квадратная поверхность оригами, узор Миура-ори, с граничной вершиной xi вдоль фронта роста (зеленый). ( B ) Фокусировка на xi, показанная с точки зрения, отличной от точки обзора A , смежных с ней вершин фронта роста xi−1 и xi+1. Два расчетных угла вдоль границы θi,3 и θi,4 показаны синим цветом, а угол в пространстве βi между двумя краями фронта роста ei и ei+1 выделен зеленым цветом.
( C ) Плоскость действия для нового расчетного угла θi,1 определяется углом закрылка αi (красный), который проходит от грани βi по часовой стрелке вокруг ei.( D ) Когда θi,1 (желтый) проходит через свою плоскость действия, он определяет возможные направления роста ri и θi,2 (пунктир), угол между ri и ei+1. Они должны удовлетворять θi,1+θi,2=ki ( D , Вставка ) для создания развертываемой вершины xi. ( E ) Это ограничение дает эллипс γi сферических дуг θi,1 и θi,2, который образует замкнутый контур вокруг линии, содержащей ei. Значение θi,1, удовлетворяющее ограничению, определяется уникальным пересечением его плоскости действия и γi, поэтому αi параметризует γi.( F ) Угол вторичного закрылка αi′ в точке xi проходит от стороны βi по часовой стрелке вокруг ei+1 и определяется αi. Угол закрылка αi+1 в точке xi+1 проходит от поверхности βi+1 по часовой стрелке вокруг ei+1 до той же плоскости, что измеряется αi’. ( G ) Два соседних направления роста ri и ri+1 должны быть компланарными, поэтому ri+1 определяется пересечением этой плоскости и γi+1; таким образом, αi+1 параметризует γi+1.
 Обозначим два проектных угла границы, падающие на xi на существующей поверхности, через θi,3 и θi,4, а угол в пространстве в точке xi вдоль фронта роста обозначим через βi=∠{−ei,ei+1}∈(0 ,π) (), где ei=xi−xi−1 и ei+1=xi+1−xi. Чтобы получить новый вектор направления ребра ri, который задает направление внутреннего ребра [xi,xi′] на расширенной четырехугольной поверхности оригами, пусть αi∈[0,2π) будет левосторонним углом закрылка вокруг ei от плоскость βi в плоскость нового четырехугольника, содержащего ri и ei (). Заметим, что одновершинное оригами в точке xi удовлетворяет условию развертываемости суммы локальных углов
Обозначим два проектных угла границы, падающие на xi на существующей поверхности, через θi,3 и θi,4, а угол в пространстве в точке xi вдоль фронта роста обозначим через βi=∠{−ei,ei+1}∈(0 ,π) (), где ei=xi−xi−1 и ei+1=xi+1−xi. Чтобы получить новый вектор направления ребра ri, который задает направление внутреннего ребра [xi,xi′] на расширенной четырехугольной поверхности оригами, пусть αi∈[0,2π) будет левосторонним углом закрылка вокруг ei от плоскость βi в плоскость нового четырехугольника, содержащего ri и ei (). Заметим, что одновершинное оригами в точке xi удовлетворяет условию развертываемости суммы локальных углов ( C ) Плоскость действия для нового расчетного угла θi,1 определяется углом закрылка αi (красный), который проходит от грани βi по часовой стрелке вокруг ei.( D ) Когда θi,1 (желтый) проходит через свою плоскость действия, он определяет возможные направления роста ri и θi,2 (пунктир), угол между ri и ei+1. Они должны удовлетворять θi,1+θi,2=ki ( D , Вставка ) для создания развертываемой вершины xi. ( E ) Это ограничение дает эллипс γi сферических дуг θi,1 и θi,2, который образует замкнутый контур вокруг линии, содержащей ei. Значение θi,1, удовлетворяющее ограничению, определяется уникальным пересечением его плоскости действия и γi, поэтому αi параметризует γi.( F ) Угол вторичного закрылка αi′ в точке xi проходит от стороны βi по часовой стрелке вокруг ei+1 и определяется αi. Угол закрылка αi+1 в точке xi+1 проходит от поверхности βi+1 по часовой стрелке вокруг ei+1 до той же плоскости, что измеряется αi’. ( G ) Два соседних направления роста ri и ri+1 должны быть компланарными, поэтому ri+1 определяется пересечением этой плоскости и γi+1; таким образом, αi+1 параметризует γi+1.
( C ) Плоскость действия для нового расчетного угла θi,1 определяется углом закрылка αi (красный), который проходит от грани βi по часовой стрелке вокруг ei.( D ) Когда θi,1 (желтый) проходит через свою плоскость действия, он определяет возможные направления роста ri и θi,2 (пунктир), угол между ri и ei+1. Они должны удовлетворять θi,1+θi,2=ki ( D , Вставка ) для создания развертываемой вершины xi. ( E ) Это ограничение дает эллипс γi сферических дуг θi,1 и θi,2, который образует замкнутый контур вокруг линии, содержащей ei. Значение θi,1, удовлетворяющее ограничению, определяется уникальным пересечением его плоскости действия и γi, поэтому αi параметризует γi.( F ) Угол вторичного закрылка αi′ в точке xi проходит от стороны βi по часовой стрелке вокруг ei+1 и определяется αi. Угол закрылка αi+1 в точке xi+1 проходит от поверхности βi+1 по часовой стрелке вокруг ei+1 до той же плоскости, что измеряется αi’. ( G ) Два соседних направления роста ri и ri+1 должны быть компланарными, поэтому ri+1 определяется пересечением этой плоскости и γi+1; таким образом, αi+1 параметризует γi+1.
, где θi,1=∠{−ei,ri}∈(0,π) и θi,2=∠{ei+1,ri}∈(0,π) — два новых проектных угла, подразумеваемых ри ().Кроме того, θi,1, θi,2 и βi образуют сферический треугольник с αi внутренним сферическим углом, противоположным θi,2, так что сферический закон косинусов дает
cosθi,2=cosθi,1cosβi+sinθi ,1sinβicosαi.
[2]
Решение уравнений. 1 и 2 и 2 для θi, 1, θi, 2 Выход
θi, 1 = Tan-1Coski-Cosβisinβicosαi-Sinei, θi, 1 ≠ π / 2,
[3]
, где ki=2π−θi,3−θi,4, количество углового материала, необходимое для обеспечения возможности развертывания.Если θi,1=π/2, мы имеем cosθi,2=sinβicosαi, что дает единственное решение, если βi≠0,π и βi
Затем можно получить новое поперечное направление кромки ri, используя θi,1 и θi,2 ().Ключевая геометрическая интуиция и альтернативное доказательство существования и единственности одновершинных решений состоит в том, чтобы заметить, что ki определяет эллипс γi из сферических дуг θi,1,θi,2, удовлетворяющий уравнению. 1 с фокусами, заданными −ei и ei+1. Для любого угла закрылка αi сумма θi,1+θi,2=βi, когда θi,1=0, и θi,1+θi,2=2π−βi, когда θi,1=π, и сумма θi,1+θi, 2 положительно монотонна на интервале θi,1∈[0,π], вообще говоря. Кроме того, неравенство сферического треугольника ограничивает βi<θi,3+θi,4<2π−βi в общем случае, поэтому независимо от того, какой угол закрылка выбран или унаследован от соседней вершины, единственное решение уравнений. 1 и 2 должны существовать, а угол закрылка αi параметризирует эллипс γi. См. Приложение SI , раздел S1 и фильм S1 для получения дополнительной информации и обсуждения.2.
Построение смежных вершин: Покажем теперь, что новые направления ребер ri+1, ri−1 в точках xi+1, xi−1 также однозначно определяются одним углом закрылка αi. Без ограничения общности рассмотрим получение ri+1 при заданном ri (). Обозначим αi′ как левосторонний угол относительно ei+1 от плоскости βi до плоскости нового четырехугольника, содержащего θi,2.Снова обращаясь к сферическому треугольнику, образованному θi,1, θi,2 и βi, сферические законы синусов и косинусов дают
 Затем можно получить новое поперечное направление кромки ri, используя θi,1 и θi,2 ().Ключевая геометрическая интуиция и альтернативное доказательство существования и единственности одновершинных решений состоит в том, чтобы заметить, что ki определяет эллипс γi из сферических дуг θi,1,θi,2, удовлетворяющий уравнению. 1 с фокусами, заданными −ei и ei+1. Для любого угла закрылка αi сумма θi,1+θi,2=βi, когда θi,1=0, и θi,1+θi,2=2π−βi, когда θi,1=π, и сумма θi,1+θi, 2 положительно монотонна на интервале θi,1∈[0,π], вообще говоря. Кроме того, неравенство сферического треугольника ограничивает βi<θi,3+θi,4<2π−βi в общем случае, поэтому независимо от того, какой угол закрылка выбран или унаследован от соседней вершины, единственное решение уравнений. 1 и 2 должны существовать, а угол закрылка αi параметризирует эллипс γi. См. Приложение SI , раздел S1 и фильм S1 для получения дополнительной информации и обсуждения.
Затем можно получить новое поперечное направление кромки ri, используя θi,1 и θi,2 ().Ключевая геометрическая интуиция и альтернативное доказательство существования и единственности одновершинных решений состоит в том, чтобы заметить, что ki определяет эллипс γi из сферических дуг θi,1,θi,2, удовлетворяющий уравнению. 1 с фокусами, заданными −ei и ei+1. Для любого угла закрылка αi сумма θi,1+θi,2=βi, когда θi,1=0, и θi,1+θi,2=2π−βi, когда θi,1=π, и сумма θi,1+θi, 2 положительно монотонна на интервале θi,1∈[0,π], вообще говоря. Кроме того, неравенство сферического треугольника ограничивает βi<θi,3+θi,4<2π−βi в общем случае, поэтому независимо от того, какой угол закрылка выбран или унаследован от соседней вершины, единственное решение уравнений. 1 и 2 должны существовать, а угол закрылка αi параметризирует эллипс γi. См. Приложение SI , раздел S1 и фильм S1 для получения дополнительной информации и обсуждения.
[5]
cosαi′=cosθi,1(αi)−cosθi,2(αi)cosβisinθi,2(αi)sinβi,
[6]
с единственным решением αi′∈[0 ,2π). Поскольку θi,1 и θi,2 являются функциями αi, αi′ также является функцией αi. Заметим, что αi′ и αi+1 измеряются вокруг общей оси и, таким образом, связаны сдвигом левостороннего угла τi от грани βi к грани βi+1.Это дает передаточную функцию угла закрылка gi:[0,2π)→[0,2π):
αi+1=gi(αi)=mod(αi′(αi)−τi,2π),
[7]
при измерении в левой ориентации относительно ei+1, начиная с плоскости βi.
Легко видеть, что g биективна; следовательно, ri+1 однозначно определяется αi, а γi и γi+1 параметризуются αi (). Аналогичный аргумент применим для ri−1. Для геометрической интуиции заметьте, что существуют биекции между точками на γi и полуплоскостями вокруг ei и ei+1.3.
Рост всего фронта: Наконец, для установления биекции между углами закрылков αi и αj при произвольных i,j, где i
 Легко видеть, что g биективна; следовательно, ri+1 однозначно определяется αi, а γi и γi+1 параметризуются αi (). Аналогичный аргумент применим для ri−1. Для геометрической интуиции заметьте, что существуют биекции между точками на γi и полуплоскостями вокруг ei и ei+1.
Легко видеть, что g биективна; следовательно, ri+1 однозначно определяется αi, а γi и γi+1 параметризуются αi (). Аналогичный аргумент применим для ri−1. Для геометрической интуиции заметьте, что существуют биекции между точками на γi и полуплоскостями вокруг ei и ei+1.αj=fi,j(αi)=gj(gj−1(gj−2(⋯gi(αi)))).
[8]
Поскольку каждая передаточная функция биективна, их композиция также биективна. Поэтому все новые направления внутренних краев вдоль всего фронта роста параметризуются одним углом закрылка αi.■
Следствие. Дана общая кривая C , дискретизированный m+1 вершин xi∈R3,i=0,…,m и м ребра ei=xi−xi−1,i=1,…,m, с углами βi=∠{−ei,ei+1}∈(0,π),i=1,…,m−1, пространство плоских паттернов, которые складываются в С это м- мерный .
Доказательство: Рассмотрим присвоение ki∈(βi,2π−βi),i=1,…,m−1 внутренним вершинам C.В приведенном выше доказательстве оригами C является фронтом роста, а ki задается существующей поверхностью оригами. Для дискретной кривой ki можно выбрать произвольно, чтобы определить одномерный набор направлений складок ri∈R3,i=1,…,m−1, которые задают развертку C в плоскость. ■
Это доказательство сразу же предлагает эффективный геометрический алгоритм для проектирования общих четырехъядерных поверхностей оригами. Для существующего правильного четырехъядерного оригами (семени) с фронтом роста, обозначенным полосой из m граничных четырехугольников (), мы отмечаем, что новая полоса из m четырехугольников имеет 3 (m + 1) степеней свободы в R3 при условии m планарности и m− 1 проектные угловые ограничения.Если мы добавим новую полосу к границе с m четырехугольниками, у нас будет в общей сложности m + 4 степени свободы для определения геометрии новой полосы: один угол створки для определения внутренних проектных углов и направлений краев, два проектных угла границы в концы полосы и m+1 длин ребер. Таким образом, в то время как наша основная теорема устанавливает пространство проектирования общего четырехъядерного оригами, следующий геометрический алгоритм позволяет нам исследовать этот ландшафт аддитивно, удовлетворяя ограничениям возможности разработки путем построения по пути.Новая совместимая полоса из m четырехугольников создается с помощью следующих шагов.
Таким образом, в то время как наша основная теорема устанавливает пространство проектирования общего четырехъядерного оригами, следующий геометрический алгоритм позволяет нам исследовать этот ландшафт аддитивно, удовлетворяя ограничениям возможности разработки путем построения по пути.Новая совместимая полоса из m четырехугольников создается с помощью следующих шагов.
1.
Начните с любого i∈{1,2,…,m−1} и выберите угол закрылка αi, связанный с фронтом роста ei (одна степень свободы) ().
2.
Распространить выбор αi вдоль фронта роста от xi до x1 и xm−1 путем итеративного решения для θi,1,θi,2, вращения ri, вычисления αi−1,αi+1 и перемещения к следующей вершине ().
3.
Выбрать проектные углы границы θ0,2,θm,1∈(0,π) и повернуть r0 и rm в положение (две степени свободы) ().
4.
Вычислить новые границы длины ребра и выбрать lj для всех j (m+1 степеней свободы). Границы задаются наблюдением, что новые ребра, обращенные наружу, в каждом новом четырехугольнике не могут пересекаться друг с другом, что происходит, когда сумма двух внутренних углов нового четырехугольника меньше π ( SI Приложение , раздел S3).
5.
Вычислить новые положения вершин, учитывая rj и lj для всех j.
6.
Повторите описанные выше шаги на любом граничном фронте, чтобы вырастить новые полосы.

Аддитивный алгоритм. ( A ) Чтобы вырастить существующую модель сложенного четырехугольника оригами на границе, состоящей из m четырехугольников, необходимо разместить в пространстве m+1 новых вершин с учетом m ограничений плоскостности (штриховые квадраты) и m−1 ограничений суммы углов (штриховые кружки) ), всего 3 (m + 1) − (2m − 1) = m + 4 степеней свободы, в общем случае. ( B ) Построение аддитивной полосы начинается с выбора плоскости, связанной с любой из четырехугольных граней в новой полосе (одна степень свободы). ( C ) Последовательные одновершинные системы распространяют этот выбор угла створки вниз на оставшуюся часть полосы, однозначно определяя ориентацию в пространстве всех четырехугольных граней в новой полосе.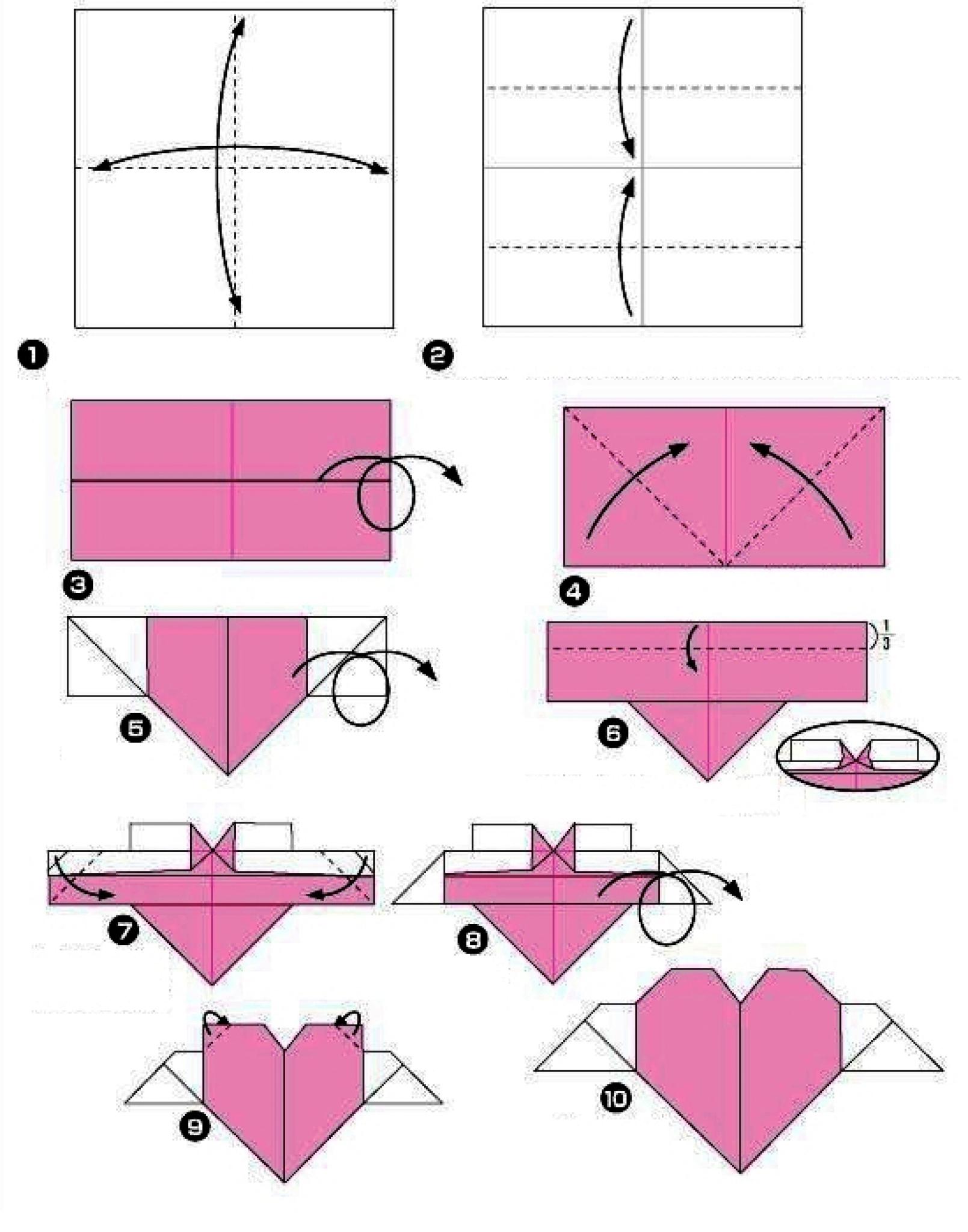 ( D ) Направления кромок в конечных точках полосы можно свободно выбирать в соответствующих плоскостях (две степени свободы), а всем поперечным кромкам в новой полосе можно присвоить длину (m+1 степень свободы) для всего m+4 степеней свободы.
( D ) Направления кромок в конечных точках полосы можно свободно выбирать в соответствующих плоскостях (две степени свободы), а всем поперечным кромкам в новой полосе можно присвоить длину (m+1 степень свободы) для всего m+4 степеней свободы.
Алгоритм также применим к дискретным кривым, не связанным с существующей складчатой поверхностью посредством следствия. В этом случае мы можем спроектировать форму развития кривой, выбрав значения k на шаге 2, а не вычисляя их по существующей поверхности.
Установив общие связи от одиночных вершин до четырех полосок и поверхностей оригами, теперь мы более подробно анализируем параметризацию угла закрылка на каждом уровне этой иерархии. Расчетное пространство фронта роста пары складчатых граней, прото-одновершинного оригами, полностью описывается парой скаляров β, угол в пространстве, образованный фронтом роста, и k, параметр формы γ — т. е. количество углового материала, необходимого для удовлетворения развертываемости (). Общее одновершинное оригами может быть построено внутри треугольной области 0≤β≤π и β≤k≤2π−β с сингулярными конфигурациями на границах, заданными равенством ().Изменение α от нуля до 2π параметризует эллипс таким образом, что θ1(α=0)=(k+β)/2 и θ1(α=π)=(k−β)/2, а новые направления ребер r(α) стремятся группироваться в пространстве вокруг направлений фронта роста, где dθ1/dα имеет меньшую величину. Специальные одновершинные оригами (40) восстанавливаются путем определения углов их закрылков (см. формулы SI, Приложение ). Три из этих типов вершин задаются перестановкой уравнения. 3 и вставка желаемого значения для нового расчетного угла: решение для продолжения αcon, где новую пару четырехугольников можно прикрепить без создания новой складки; плоскоскладное решение αff, где вершину можно полностью сложить так, что все грани компланарны; и αeq, что создает равные новые конструктивные углы.Каждое из них допускает два решения, связанных отражением над плоскостью фронта роста, что восстанавливает двойственность, отмеченную в [2].
Общее одновершинное оригами может быть построено внутри треугольной области 0≤β≤π и β≤k≤2π−β с сингулярными конфигурациями на границах, заданными равенством ().Изменение α от нуля до 2π параметризует эллипс таким образом, что θ1(α=0)=(k+β)/2 и θ1(α=π)=(k−β)/2, а новые направления ребер r(α) стремятся группироваться в пространстве вокруг направлений фронта роста, где dθ1/dα имеет меньшую величину. Специальные одновершинные оригами (40) восстанавливаются путем определения углов их закрылков (см. формулы SI, Приложение ). Три из этих типов вершин задаются перестановкой уравнения. 3 и вставка желаемого значения для нового расчетного угла: решение для продолжения αcon, где новую пару четырехугольников можно прикрепить без создания новой складки; плоскоскладное решение αff, где вершину можно полностью сложить так, что все грани компланарны; и αeq, что создает равные новые конструктивные углы.Каждое из них допускает два решения, связанных отражением над плоскостью фронта роста, что восстанавливает двойственность, отмеченную в [2]. 5. Еще две специальные вершины обозначаются αll и αlr, которые образуют заблокированные конфигурации с левой парой граней (θ1,θ4) и правой парой (θ2,θ3), свернутыми до компланарности соответственно. Вершина тривиально блокируется компланарными (θ1,θ2) гранями при α=0,π. Угол продолжения лоскута, который не создает новую складку вдоль фронта роста, и угол заблокированного левого лоскута связаны соотношением αcon=mod(αll+π,2π).Отметим также, что самопересечение будет иметь место для углов закрылков в промежутках между плоскостью β и ближайшим нетривиальным запертым углом закрылков. Примечательно, что в нашей конструкции отсутствуют углы сгиба, которые можно восстановить на краю фронта роста ei с помощью ϕi,p=αi,ll−αi−π.
5. Еще две специальные вершины обозначаются αll и αlr, которые образуют заблокированные конфигурации с левой парой граней (θ1,θ4) и правой парой (θ2,θ3), свернутыми до компланарности соответственно. Вершина тривиально блокируется компланарными (θ1,θ2) гранями при α=0,π. Угол продолжения лоскута, который не создает новую складку вдоль фронта роста, и угол заблокированного левого лоскута связаны соотношением αcon=mod(αll+π,2π).Отметим также, что самопересечение будет иметь место для углов закрылков в промежутках между плоскостью β и ближайшим нетривиальным запертым углом закрылков. Примечательно, что в нашей конструкции отсутствуют углы сгиба, которые можно восстановить на краю фронта роста ei с помощью ϕi,p=αi,ll−αi−π.
Дизайн вершин и полос. ( A ) Показана пара сложенных четырехугольников и их одновершинный фронт роста с β = π/2 и сферический эллипс, заданный k = 5π/4, а также два других слабых эллипса, заданных k = π, 3π /4, что было бы дано другими существующими расчетными углами, чем те, которые показаны. Направления роста особых вершин и интервалы самопересечения восстанавливаются путем определения их углов крыльев, три из которых (αcon, αeq, αff) имеют два решения, полученные путем отражения по плоскости β. ( B , Верхний ) Допустимая область для β и k в одной вершине. Типичные общие вершины фронта роста (красные, зеленые и синие точки) попадают внутрь этой области. Первый расчетный угол θ1 ограничен сверху величиной (k+β)/2 при α=0 и снизу величиной (k−β)/2 при α=π, а в зазоре показаны поверхности с постоянным α.Подметание α∈[0,2π) дает возможные значения для θ1 ( B , Lower ), симметричные относительно α=π для трех цветных точек, указанных выше. ( C ) Сложенная четверная полоса с двумя совместимыми направлениями роста (продолжение, при котором не создается новая складка, и сложенная конфигурация), выбранная из одномерного пространства совместимых конструкций полос, параметризованная ориентацией в пространстве первой новой лицо. ( D ) Половина новых углов внутреннего оформления θi,1 в новой полосе ( Верх ), углы сгиба поперек фронта роста ϕi,t ( Середина ) и параллельно фронту роста ϕi,p ( Нижняя часть ) показаны как функции угла закрылка α1.
Направления роста особых вершин и интервалы самопересечения восстанавливаются путем определения их углов крыльев, три из которых (αcon, αeq, αff) имеют два решения, полученные путем отражения по плоскости β. ( B , Верхний ) Допустимая область для β и k в одной вершине. Типичные общие вершины фронта роста (красные, зеленые и синие точки) попадают внутрь этой области. Первый расчетный угол θ1 ограничен сверху величиной (k+β)/2 при α=0 и снизу величиной (k−β)/2 при α=π, а в зазоре показаны поверхности с постоянным α.Подметание α∈[0,2π) дает возможные значения для θ1 ( B , Lower ), симметричные относительно α=π для трех цветных точек, указанных выше. ( C ) Сложенная четверная полоса с двумя совместимыми направлениями роста (продолжение, при котором не создается новая складка, и сложенная конфигурация), выбранная из одномерного пространства совместимых конструкций полос, параметризованная ориентацией в пространстве первой новой лицо. ( D ) Половина новых углов внутреннего оформления θi,1 в новой полосе ( Верх ), углы сгиба поперек фронта роста ϕi,t ( Середина ) и параллельно фронту роста ϕi,p ( Нижняя часть ) показаны как функции угла закрылка α1. Характерные одновершинные кривые, связанные с x1, выделены жирным шрифтом, а кривые, связанные с другими вершинами (светло-черные), отличаются от характерных одновершинных кривых нелокальностью.
Характерные одновершинные кривые, связанные с x1, выделены жирным шрифтом, а кривые, связанные с другими вершинами (светло-черные), отличаются от характерных одновершинных кривых нелокальностью.
Двигаясь вверх по шкале, мы исследуем взаимосвязь между углом закрылка и дизайном полосы. Специальные одновершинные решения не обязательно могут быть применены во всех точках общего фронта роста поверхности, поскольку пространство направлений роста является одномерным. Например, для создания набора плоскоскладываемых складок фронта роста требуются дополнительные симметрии.Исключением является αi,con, единственное значение угла закрылка, которое дает тривиальное направление роста для всего фронта. В , мы иллюстрируем общую свернутую четверную полосу и две ее совместимые полосы, тривиальное решение продолжения и нетривиальное свернутое решение. Новые конструктивные углы θi,1, углы сгиба, поперечные фронту роста ϕi,t и параллельные фронту роста ϕi,p в новой полосе, показаны как функции угла закрылка α1 в . Углы сгиба, параллельные фронту роста ϕi,p, одновременно равны нулю при α1,con и отличны от нуля в противном случае, в то время как углы сгиба поперек фронта роста ϕi,t вообще никогда не равны нулю.См. Приложение SI , раздел S2 и фильмы S2 и S3 для получения более подробной информации.
Углы сгиба, параллельные фронту роста ϕi,p, одновременно равны нулю при α1,con и отличны от нуля в противном случае, в то время как углы сгиба поперек фронта роста ϕi,t вообще никогда не равны нулю.См. Приложение SI , раздел S2 и фильмы S2 и S3 для получения более подробной информации.
Чтобы продемонстрировать возможности нашего аддитивного подхода, мы теперь применяем его в рамках обратного проектирования для построения упорядоченных и неупорядоченных четырехъядерных типологий оригами с прямыми и изогнутыми складками. В отличие от предыдущей работы (32), наш аддитивный подход не требует решения большой многомерной задачи оптимизации всей структуры. Вместо этого требуется только выбор из доступных степеней свободы для каждой полосы, которые отображают все пространство совместимых дизайнов и, следовательно, являются более выполнимыми с точки зрения вычислений и геометрически полными.Эти варианты зависят от приложения и могут быть случайными, интерактивными или основываться на некоторых критериях оптимизации.
В качестве первого примера мы рассмотрим аппроксимацию двойной криволинейной целевой поверхности с помощью обобщенной мозаики Миура-ори. Для заданной гладкой целевой поверхности, которую мы хотим аппроксимировать, рассмотрим две граничные поверхности, смещенные в нормальном направлении от целевой поверхности (верхнюю и нижнюю границы), и построим в их междоузлиях простую однократно гофрированную полосу с одной стороной полоски лежащий на верхней поверхности и одной стороной на нижней поверхности (см. Приложение SI , раздел S4A для получения более подробной информации).Затем, применяя наш аддитивный алгоритм, мы добавляем полоски по обеим сторонам семени (и продолжая на участке роста), которые примерно отражают поверхность оригами вперед и назад между верхней и нижней границами цели, вызывая дополнительную гофрировку в поперечном направлении для что из гофры в семени. показывает обобщенную многослойную структуру Миура-ори постоянной толщины с высоким разрешением, полученную с помощью нашего подхода, которая аппроксимирует ландшафт смешанной кривизны, который было бы очень трудно получить с использованием современных методов. В качестве второго примера, используя другую настройку выбора степени свободы без эталонной целевой поверхности, мы выращиваем коническую затравку с рядом прямых гребней с четырехкратной симметрией через грани, которые создаются отражениями туда и обратно между парой вращающихся плоскостей, как показано на . В качестве нашего третьего примера мы обратимся к проектированию поверхностей, которые имеют кривые складки, складки, которые аппроксимируют гладкую пространственную кривую с ненулевой кривизной и кручением (5). показывает искривленную версию концентрической круглой башни (41) Дэвида Хаффмана , полученную нашим методом, в котором используется аналогичная установка выбора степени свободы, но начинается с сегмента конуса с высоким разрешением в качестве затравки внутреннего кольца, а затем с постепенно увеличивающимся наклоном. конические кольца добавлены поперечно (см. SI Приложение , раздел S4B для получения более подробной информации по обеим этим моделям).показана другая модель криволинейной складки, которая использует следствие для создания затравки из гофрированной дискретной плоской параболы и продолжает добавлять новые полосы с постоянными углами сгиба и длинами ребер, растущими в направлении вдоль складок (см.
В качестве второго примера, используя другую настройку выбора степени свободы без эталонной целевой поверхности, мы выращиваем коническую затравку с рядом прямых гребней с четырехкратной симметрией через грани, которые создаются отражениями туда и обратно между парой вращающихся плоскостей, как показано на . В качестве нашего третьего примера мы обратимся к проектированию поверхностей, которые имеют кривые складки, складки, которые аппроксимируют гладкую пространственную кривую с ненулевой кривизной и кручением (5). показывает искривленную версию концентрической круглой башни (41) Дэвида Хаффмана , полученную нашим методом, в котором используется аналогичная установка выбора степени свободы, но начинается с сегмента конуса с высоким разрешением в качестве затравки внутреннего кольца, а затем с постепенно увеличивающимся наклоном. конические кольца добавлены поперечно (см. SI Приложение , раздел S4B для получения более подробной информации по обеим этим моделям).показана другая модель криволинейной складки, которая использует следствие для создания затравки из гофрированной дискретной плоской параболы и продолжает добавлять новые полосы с постоянными углами сгиба и длинами ребер, растущими в направлении вдоль складок (см.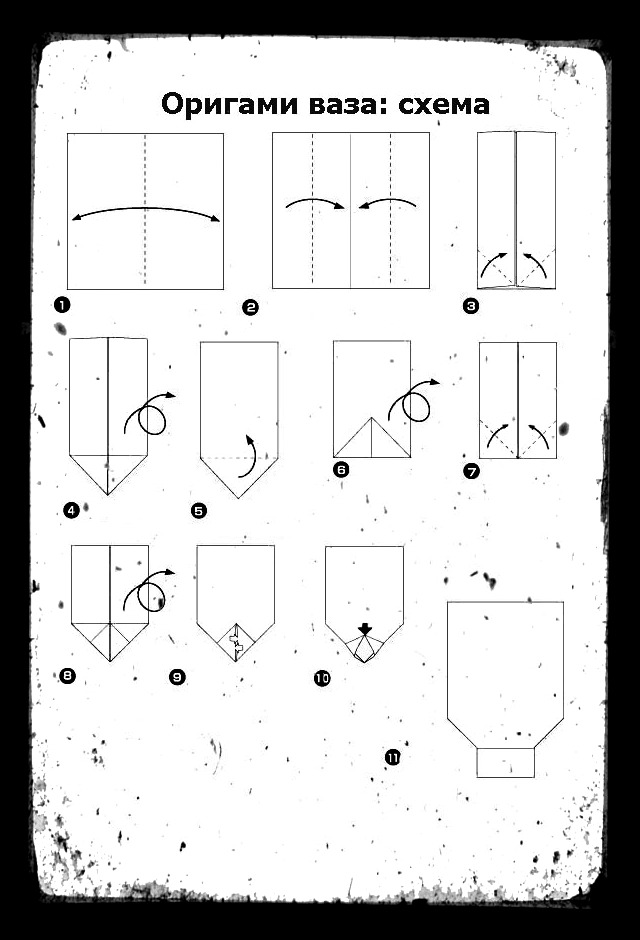 SI Приложение , раздел S4C Больше подробностей). В качестве нашего последнего примера поверхности мы используем наш подход для создания неупорядоченной, скомканной поверхности, изометричной плоскости, опять же структуру, которую было бы очень трудно получить с использованием современных методов. Для каждого шага построения полосы в аддитивном алгоритме угол лоскута и длина края могут быть выбраны случайным образом, что приводит к смятому листу, который не соответствует заданному шаблону MV ().Для моделирования физически реализуемой смятой геометрии мы выбрали углы закрылков в соответствии с границами самопересечения, заданными специальными вершинными решениями по всему фронту, так что рост листа является локально самоизбегающим блужданием (см. SI Приложение ). , раздел S4D для более подробной информации).
SI Приложение , раздел S4C Больше подробностей). В качестве нашего последнего примера поверхности мы используем наш подход для создания неупорядоченной, скомканной поверхности, изометричной плоскости, опять же структуру, которую было бы очень трудно получить с использованием современных методов. Для каждого шага построения полосы в аддитивном алгоритме угол лоскута и длина края могут быть выбраны случайным образом, что приводит к смятому листу, который не соответствует заданному шаблону MV ().Для моделирования физически реализуемой смятой геометрии мы выбрали углы закрылков в соответствии с границами самопересечения, заданными специальными вершинными решениями по всему фронту, так что рост листа является локально самоизбегающим блужданием (см. SI Приложение ). , раздел S4D для более подробной информации).
Аддитивное проектирование прямых, изогнутых, упорядоченных и беспорядочных оригами. ( A ) Обобщенная мозаика Миура-ори, подходящая к целевой поверхности со смешанной гауссовой кривизной.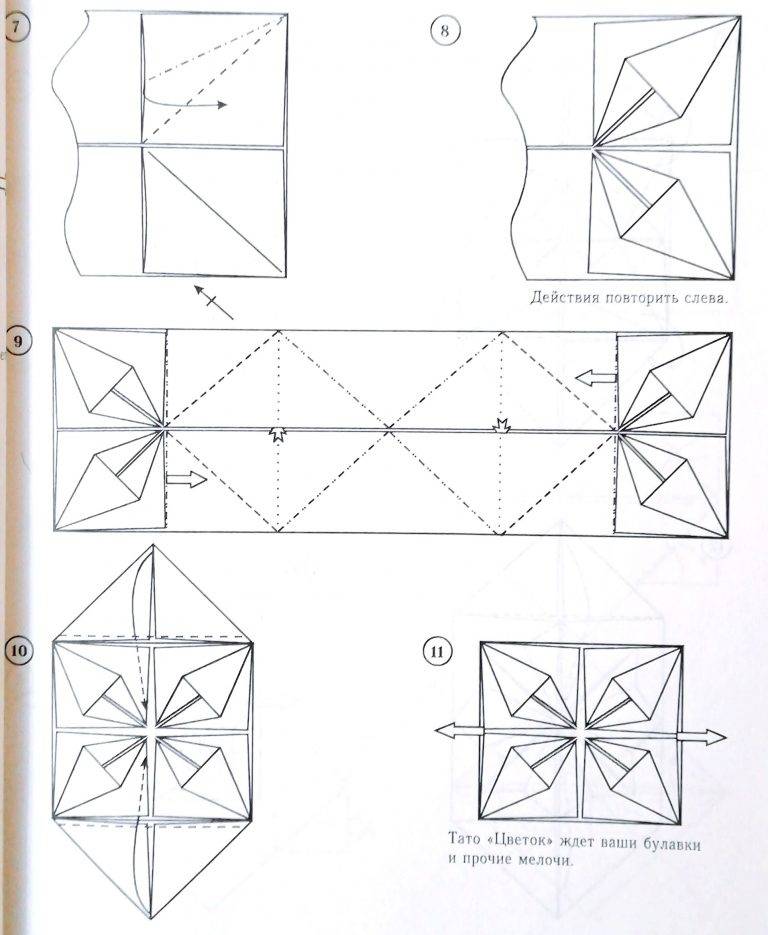 Нижняя и верхняя граничные поверхности смещаются нормально от целевой поверхности, и между ними инициализируется затравочная полоса четырехугольников с одним фронтом роста на каждой поверхности. Новые полоски прикрепляются по обе стороны от семени за счет отражения фронта роста вперед и назад между граничными поверхностями в их междоузлиях. ( B ) Коническое зародыш низкого разрешения с четырехкратной симметрией растет путем отражения между промежутками двух вращающихся верхней и нижней граничных плоскостей. Новые полосы образуют замкнутые петли с перекрывающимися гранями конечных точек.( C ) Коническое семя с высоким разрешением растет путем прикрепления конусных колец с прогрессивным наклоном, чтобы воспроизвести модель изогнутой складки. Новые полосы образуют замкнутые петли с перекрывающимися гранями конечных точек. ( D ) Модель криволинейной складки вырастает из затравки, созданной путем использования следствия для присоединения полосы четырехугольников к гофрированной параболе.
Нижняя и верхняя граничные поверхности смещаются нормально от целевой поверхности, и между ними инициализируется затравочная полоса четырехугольников с одним фронтом роста на каждой поверхности. Новые полоски прикрепляются по обе стороны от семени за счет отражения фронта роста вперед и назад между граничными поверхностями в их междоузлиях. ( B ) Коническое зародыш низкого разрешения с четырехкратной симметрией растет путем отражения между промежутками двух вращающихся верхней и нижней граничных плоскостей. Новые полосы образуют замкнутые петли с перекрывающимися гранями конечных точек.( C ) Коническое семя с высоким разрешением растет путем прикрепления конусных колец с прогрессивным наклоном, чтобы воспроизвести модель изогнутой складки. Новые полосы образуют замкнутые петли с перекрывающимися гранями конечных точек. ( D ) Модель криволинейной складки вырастает из затравки, созданной путем использования следствия для присоединения полосы четырехугольников к гофрированной параболе. ( E ) Уход от посевной полосы Миура-ори с шумом, добавленным к граничному фронту роста, приводит к смятому листу. Новые полосы добавляются путем выборки углов закрылков в пределах, которые предотвращают локальное самопересечение.( F ) Броуновская лента, развитие которой приближается к круговому кольцу, создается с помощью следствия. Семена выделены желтым цветом, а стрелки указывают направление роста. Шаблон сгиба для каждой модели показан в правом нижнем углу каждого изображения. Верхние правые вставки в A–C и F вместе с небольшим квадратом рядом с шаблоном в F представляют собой увеличенные виды, чтобы выделить детали. См. Приложение SI , рис.S8 и S9 для версий шаблонов сгиба с более высоким разрешением.
( E ) Уход от посевной полосы Миура-ори с шумом, добавленным к граничному фронту роста, приводит к смятому листу. Новые полосы добавляются путем выборки углов закрылков в пределах, которые предотвращают локальное самопересечение.( F ) Броуновская лента, развитие которой приближается к круговому кольцу, создается с помощью следствия. Семена выделены желтым цветом, а стрелки указывают направление роста. Шаблон сгиба для каждой модели показан в правом нижнем углу каждого изображения. Верхние правые вставки в A–C и F вместе с небольшим квадратом рядом с шаблоном в F представляют собой увеличенные виды, чтобы выделить детали. См. Приложение SI , рис.S8 и S9 для версий шаблонов сгиба с более высоким разрешением.
Наконец, чтобы подчеркнуть гибкость следствия, мы построим четверную полосу, образующую складную связь между канонически грубой структурой, случайным блужданием в трех измерениях (3D), и канонически гладкой структурой, кругом в двух измерениях ( 2Д). показывает единственную сложенную полосу, сгенерированную путем выборки дискретного броуновского пути в 3D для формирования одной границы сложенной полосы, выбирая значения k таким образом, чтобы ее развитие падало на окружность и образовывало единую замкнутую петлю для любого выбора значения угла закрылка, и выбирая такие длины краев, чтобы другая граница полосы переходила в другую, меньшую концентрическую окружность ( Приложение SI , раздел S4E).Действительно, это следствие позволяет создавать произвольную форму как сложенных лент, так и их аналогов с узором независимо друг от друга. См. фильмы S4–S9 для 3D-анимации моделей и SI Приложение , рис. S11, S12, S14, S16 и S17 для галереи других результатов подгонки поверхности, изогнутых складок, неупорядоченных и броуновских лент, полученных с помощью нашего аддитивного подхода.
показывает единственную сложенную полосу, сгенерированную путем выборки дискретного броуновского пути в 3D для формирования одной границы сложенной полосы, выбирая значения k таким образом, чтобы ее развитие падало на окружность и образовывало единую замкнутую петлю для любого выбора значения угла закрылка, и выбирая такие длины краев, чтобы другая граница полосы переходила в другую, меньшую концентрическую окружность ( Приложение SI , раздел S4E).Действительно, это следствие позволяет создавать произвольную форму как сложенных лент, так и их аналогов с узором независимо друг от друга. См. фильмы S4–S9 для 3D-анимации моделей и SI Приложение , рис. S11, S12, S14, S16 и S17 для галереи других результатов подгонки поверхности, изогнутых складок, неупорядоченных и броуновских лент, полученных с помощью нашего аддитивного подхода.
Поскольку условие развертываемости в уравнении. 1 всегда выполняется в нашем маршевом алгоритме, все созданные им физические модели, которые не самопересекаются, могут трансформироваться из двухмерного плоского состояния в изометрическое трехмерное свернутое состояние, как правило, посредством энергетического ландшафта, включающего геометрическую фрустрацию (см. SI Приложение , раздел S5 и рис.S18 и Movie S10 для имитации складывания). Поскольку ландшафт зависит от геометрии шаблона складывания, для которого движения складывания не уникальны, это открывает будущие пути для программной метастабильности. Заметим также, что, заменив правую часть в уравнении 1 на K≠2π и соответствующим образом модифицируя последующие тригонометрические формулы, наш аддитивный подход обобщается на построение неевклидовых оригами (42, 43), с решениями, однозначно существующими таким же образом, когда K−θi,3−θi, 4∈(βi,2π−βi).
SI Приложение , раздел S5 и рис.S18 и Movie S10 для имитации складывания). Поскольку ландшафт зависит от геометрии шаблона складывания, для которого движения складывания не уникальны, это открывает будущие пути для программной метастабильности. Заметим также, что, заменив правую часть в уравнении 1 на K≠2π и соответствующим образом модифицируя последующие тригонометрические формулы, наш аддитивный подход обобщается на построение неевклидовых оригами (42, 43), с решениями, однозначно существующими таким же образом, когда K−θi,3−θi, 4∈(βi,2π−βi).
В целом, наше исследование обеспечивает единую основу для обратного проектирования общих развертываемых четырехъядерных шаблонов оригами и дискретных развертываемых поверхностей посредством роста. Простая теорема формирует основу маршевого алгоритма, который заменяет решение сложной задачи глобальной оптимизации масштабируемой, простой в реализации схемой эволюции складчатого фронта с ограничениями. Это взаимодействие между объемной жесткостью и граничной гибкостью, которое позволяет нам быстро создавать прототипы вычислительных конструкций упорядоченной, неупорядоченной, прямолинейной и криволинейно-сложенной геометрии, сулит существенные перспективы для достижений как в дискретной геометрии, инженерных приложениях, так и в художественных произведениях.
Группа 3D Optical Systems
ВВЕДЕНИЕПока двумерные (2D) методы нанопроизводства, такие как электронная лучевая и рентгеновская литография, наноимпринтинг, перьевая литография, и т. д., могут надежно воспроизводить элементы размером менее 20 нм, эти методы отсутствие возможности создавать неплоские узоры, т.е.е. трехмерный (3D) структуры. Использование 3-го измерения позволит устройств, чтобы обеспечить дополнительную гибкость или даже полностью новые функции. Третье измерение также помогает преодолеть связанные задача преодоления разрыва между наноразмерными характеристиками системы и макромасштаба (рис. 1).
Процесс Nanostructured Origami™ преодолевает эту трудность
использования 3-го измерения путем заимствования идей из
Японское искусство складывания бумаги или оригами.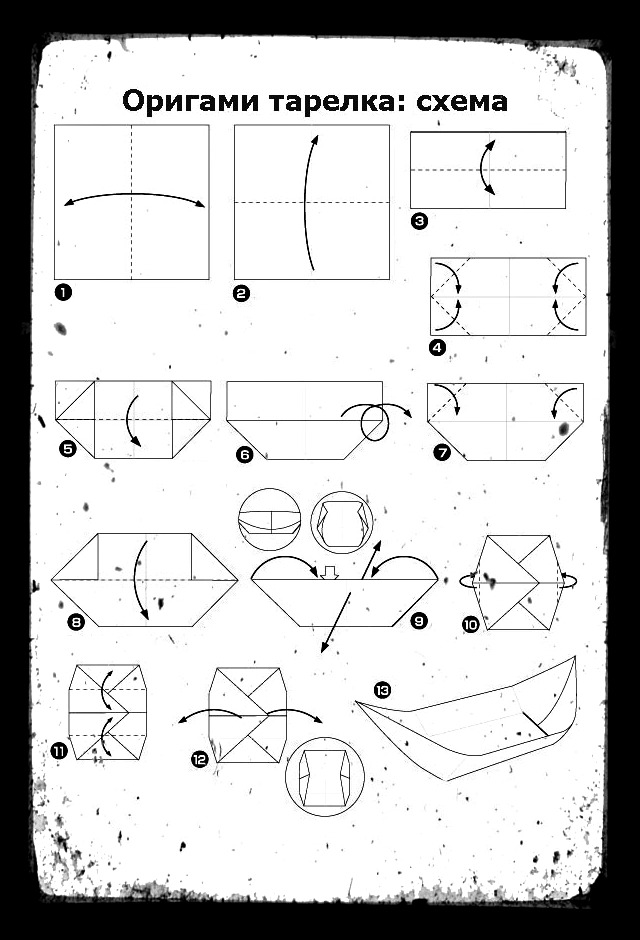 По первому паттерну
2D-мембраны с помощью обычных инструментов литографии, а затем складывание
их в произвольную компоновку, почти любая трехмерная форма может быть
сформировался. Конечно, каждая из этих складчатых мембран будет иметь
был создан на предыдущем шаге с желаемым механическим,
электрические, оптические и другие функции (рис. 2 и 3).
По первому паттерну
2D-мембраны с помощью обычных инструментов литографии, а затем складывание
их в произвольную компоновку, почти любая трехмерная форма может быть
сформировался. Конечно, каждая из этих складчатых мембран будет иметь
был создан на предыдущем шаге с желаемым механическим,
электрические, оптические и другие функции (рис. 2 и 3).
Рисунок
1 Преодоление разрыва между наномасштабом и макромасштабом.
(а)
(до н.э)
Рисунок 2 (а) Плоское изготовление. (b) Складывание мембраны.
(c) Готовое устройство.
ТОК ПРОГРЕСС СКЛАДНОЙ МЕТОДЫ
В настоящее время мы изучаем два основных метода создания оригами. складывание мембраны: срабатывание силы Лоренца и остаточное напряжение
индуцированное скручивание.
складывание мембраны: срабатывание силы Лоренца и остаточное напряжение
индуцированное скручивание.
В метод действия силы Лоренца, ток подается на сегмент, который должен складываться в присутствии магнитного поля. По закону Лоренца на него действует направленная вверх сила. внешний край сегмента, как показано на рисунке 4.
Другой
Схема складывания, которую мы исследуем, относится к нитриду кремния
как «бумага.” Области складок сначала литографически
определяется, а складывание вызывается осаждением напряженного металла
слой поверх нитрида. Мы можем сложить нитрид
слой под произвольным углом, регулируя размер складки
и управление остаточным напряжением наплавленного металла
исполнительный слой.
Рисунок 5 показано изображение, полученное с помощью сканирующего электронного микроскопа (СЭМ), слой сложен за пределы 360o.Радиус кривизны примерно 20 микрон, что примерно равно радиусу человеческого волоса.
Рисунок 6 показан вид сверху на скручивание. Верхний слой хром и выглядит белым под микроскопом. Нижний слой, силикон нитрид, имеет более темный зеленый цвет. Можно наблюдать складывание от внешних углов.
Рис. 4. Сгибание мембраны под действием силы Лоренца
Рис. 5: СЭМ-изображение 360-градусной складки.
Рис. 6: Завивка, вид сверху.
Несколько устройств были успешно изготовлены. Наш первый
устройства были изготовлены с использованием кремния в качестве конструкционного материала
закрылков и складываются на 180º с помощью вышеупомянутого
Метод срабатывания силы Лоренца.Решетки с характеристиками менее 40 нм
также были включены в изготовленные устройства. Следующий
Поколение устройств использовало полимер под названием SU-8 в качестве структурного элемента.
материал. На рис. 8 показано двухстворчатое устройство из СУ-8, которое
выскочил из подложки из-за вызванного стрессом
схема складывания упоминалась ранее. На рис. 9 показана пирамида.
структуры, которые используются для поддержания расстояния и выравнивания
между различными складчатыми слоями.На рис. 10 показано устройство
после завершения этапа складывания.
Наш первый
устройства были изготовлены с использованием кремния в качестве конструкционного материала
закрылков и складываются на 180º с помощью вышеупомянутого
Метод срабатывания силы Лоренца.Решетки с характеристиками менее 40 нм
также были включены в изготовленные устройства. Следующий
Поколение устройств использовало полимер под названием SU-8 в качестве структурного элемента.
материал. На рис. 8 показано двухстворчатое устройство из СУ-8, которое
выскочил из подложки из-за вызванного стрессом
схема складывания упоминалась ранее. На рис. 9 показана пирамида.
структуры, которые используются для поддержания расстояния и выравнивания
между различными складчатыми слоями.На рис. 10 показано устройство
после завершения этапа складывания.
Рисунок 7: Интегрированные решетки наноразмерных элементов
в складное устройство
Рис. 8: Двухстворчатое устройство после индуцированного частичного складывания
8: Двухстворчатое устройство после индуцированного частичного складывания
через остаточное напряжение.
(а)
(b)
Рисунок 9: (a) Оригами с использованием процесса SU-8.(б) Крупный план
вид на пирамиды выравнивания.
Рисунок 10: Изображение сверху двухстворчатого устройства.
Программное обеспечение
была разработана для визуализации процесса складывания и
моделирование кинематики конструкций оригами. Наша модель начинается
путем присоединения замкнутой «каркасной» конструкции к
сегменты оригами.Затем скелет разбивается на две части.
части и поступательная кинематика выполняются в той части, которая
содержит исполнительный механизм.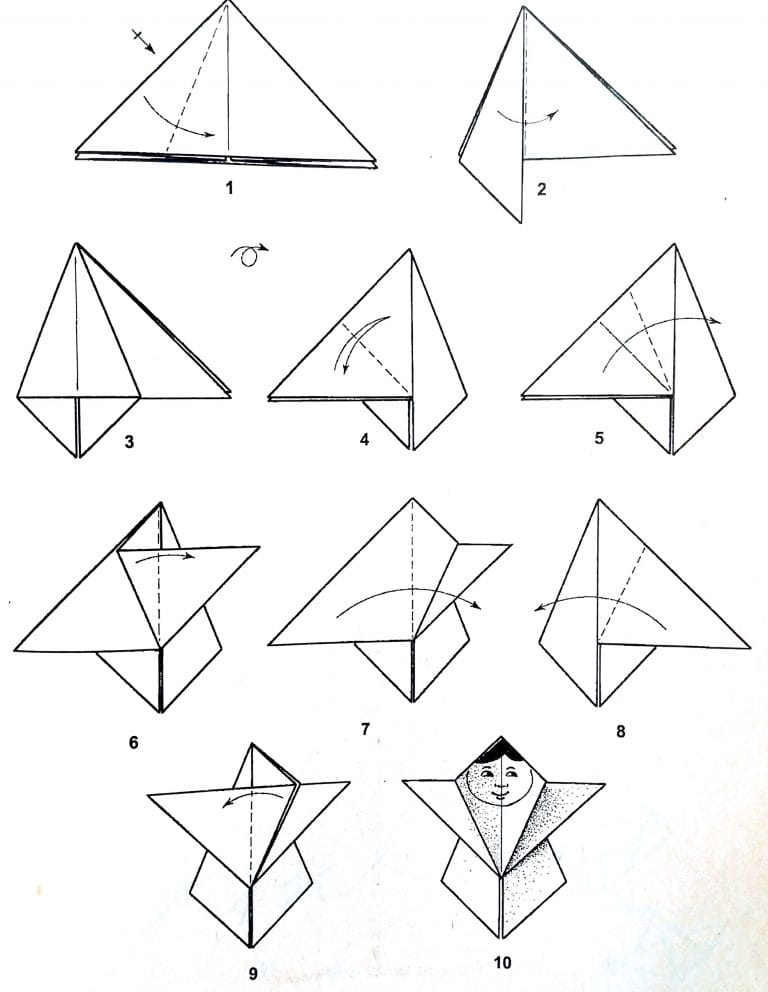 Вторая часть ограничена
по условию целостности, поэтому его движение оценивается
с помощью инверсной кинематики. Особенности инверсной кинематики
второй части свидетельствует о нарушении кинематической целостности
в заданной структуре. Кинематическая совместимость обеспечена
путем отслеживания краев сегмента (эта функция еще не
реализованы в нашем программном инструменте).Графический интерфейс разработанного
программное обеспечение показано на рис. 10, а на рис. 11 показан пример
процесса визуализации и кинематического моделирования [1].
Вторая часть ограничена
по условию целостности, поэтому его движение оценивается
с помощью инверсной кинематики. Особенности инверсной кинематики
второй части свидетельствует о нарушении кинематической целостности
в заданной структуре. Кинематическая совместимость обеспечена
путем отслеживания краев сегмента (эта функция еще не
реализованы в нашем программном инструменте).Графический интерфейс разработанного
программное обеспечение показано на рис. 10, а на рис. 11 показан пример
процесса визуализации и кинематического моделирования [1].
Рисунок
11: Программное обеспечение, разработанное для визуализации и выполнения кинематики.
моделирование процесса складывания.
Рисунок 12: Пятисегментный механизм оригами (последовательность срабатывания
показано по часовой стрелке сверху) и углы складывания двух
сегментов в ответ на активацию диагонального сустава.
Наноструктурный процесс изготовления и сборки Origami™ был разработан для 3D-нанопроизводства с использованием обычных Инструменты для двумерной литографии. Результаты, представленные выше, показывают большие надежды для этой техники, и мы уже находимся в процессе применения этот метод для изготовления новых трехмерных наноструктурированных устройства, такие как системы накопления энергии и многослойные дифракционные оптические элементы.
НОМЕР:
1. Т. Бюхнер, «Кинематика трехмерных складных структур для наноструктурного оригами», дипломная работа, Лаборатория станков и технологии производства, Рейнско-Вестфальский технический университет Ахена (2004).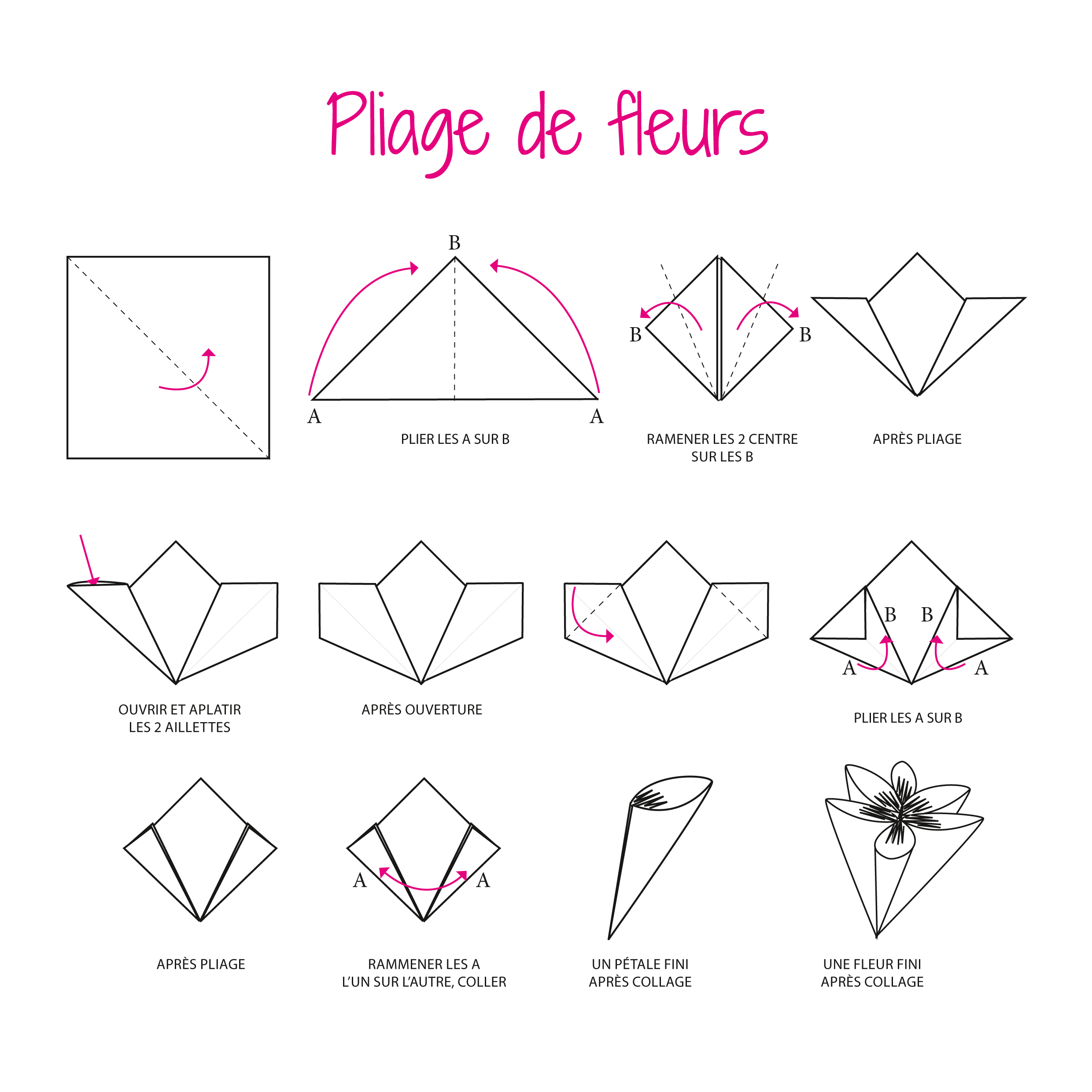
НАВЕРХ
Стабильные трехмерные иерархические каркасы с использованием подхода оригами: эффект межфазного сшивания наногибридными сборками шашлыка
https://doi.org/10.1016/j.matdes.2021.110353Получить права и содержимоеОсновные моменты
- •
Полимерные волокнистые слои, армированные нанонаполнителями, были изготовлены для создания гибких каркасов путем складывания 2D-мата в многослойные стопки с использованием подхода, вдохновленного оригами.
- •
Взаимопроникающая сеть гибридных полимерных ламелей была выращена на стыках для улучшения связи между последующими строительными блоками.
- •
Стабильность размеров контролировалась по шкале длины для получения высокостабильной иерархической геометрии пор.
Abstract
Трехмерные высокопористые архитектуры с индивидуальной гибкостью и стабильностью размеров в настоящее время являются новой областью благодаря широкому спектру технологических применений.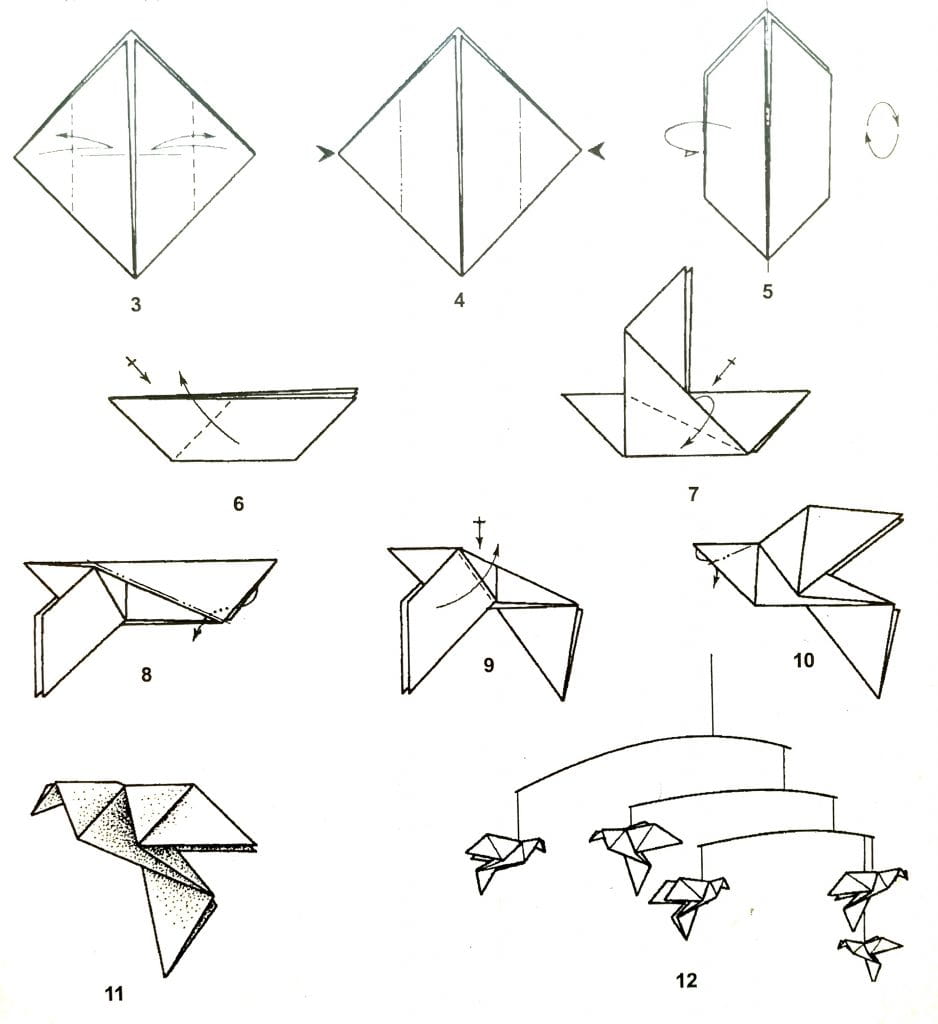 Здесь мы подчеркнули важность механической и термической стабильности строительных блоков и межфазного соединения для разработки таких каркасов. 2D электроформованные отдельно стоящие нановолокнистые маты из полиэтилентерефталата (ПЭТФ) с различной молекулярной массой были использованы для изучения свойств основных блоков основы.Сшитые сети наногибридного шашлыка (NHSK) типа поликапролактона (PCL) были выращены между штабелированным нановолокнистым матом для лучшего связывания между слоями. Углеродные нанотрубки были дополнительно введены в нановолокна ПЭТФ, а также в связующие покровные слои, чтобы понять свойства термической и структурной стабильности. Наличие различных ограниченных одномерных зародышевых поверхностей, таких как нанотрубки и нановолокна, дает нам возможность изучать взаимопроникающие ламели NHSK на разных масштабах длины.Ряд сшитых каркасных микроструктур методом оригами был исследован, чтобы подчеркнуть важность проектирования процесса для изготовления размерно-устойчивых пористых геометрий.
Здесь мы подчеркнули важность механической и термической стабильности строительных блоков и межфазного соединения для разработки таких каркасов. 2D электроформованные отдельно стоящие нановолокнистые маты из полиэтилентерефталата (ПЭТФ) с различной молекулярной массой были использованы для изучения свойств основных блоков основы.Сшитые сети наногибридного шашлыка (NHSK) типа поликапролактона (PCL) были выращены между штабелированным нановолокнистым матом для лучшего связывания между слоями. Углеродные нанотрубки были дополнительно введены в нановолокна ПЭТФ, а также в связующие покровные слои, чтобы понять свойства термической и структурной стабильности. Наличие различных ограниченных одномерных зародышевых поверхностей, таких как нанотрубки и нановолокна, дает нам возможность изучать взаимопроникающие ламели NHSK на разных масштабах длины.Ряд сшитых каркасных микроструктур методом оригами был исследован, чтобы подчеркнуть важность проектирования процесса для изготовления размерно-устойчивых пористых геометрий.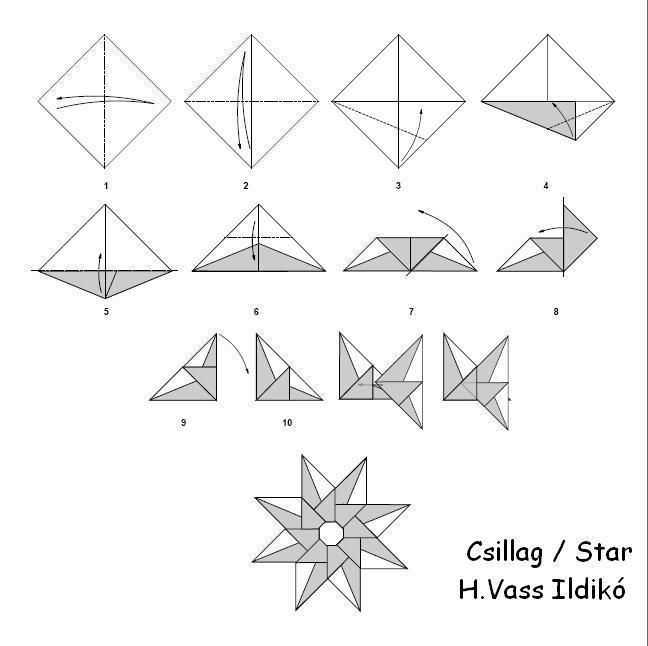 (0) Опубликовано Elsevier Ltd.
(0) Опубликовано Elsevier Ltd.
Рекомендуемые статьи
Ссылающиеся статьи
Трехмерная динамика формы одиночных клеток, ориентированная вдоль апико-базальной оси складчатого эпителия по данным флуоресцентной микроскопии
Цитирование: , Frangi AF (2021)Оригами: динамика трехмерной формы одиночных клеток, ориентированная вдоль апико-базальной оси складчатого эпителия, по данным флуоресцентной микроскопии. PLoS Comput Biol 17(11): е1009063. https://дои.org/10.1371/journal.pcbi.1009063
Редактор: Дина Шнейдман-Духовны, Еврейский университет в Иерусалиме, ИЗРАИЛЬ
Поступила в редакцию: 5 мая 2021 г.; Принято: 13 октября 2021 г .; Опубликовано: 1 ноября 2021 г.
Авторское право: © 2021 Mendonca et al. Это статья с открытым доступом, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.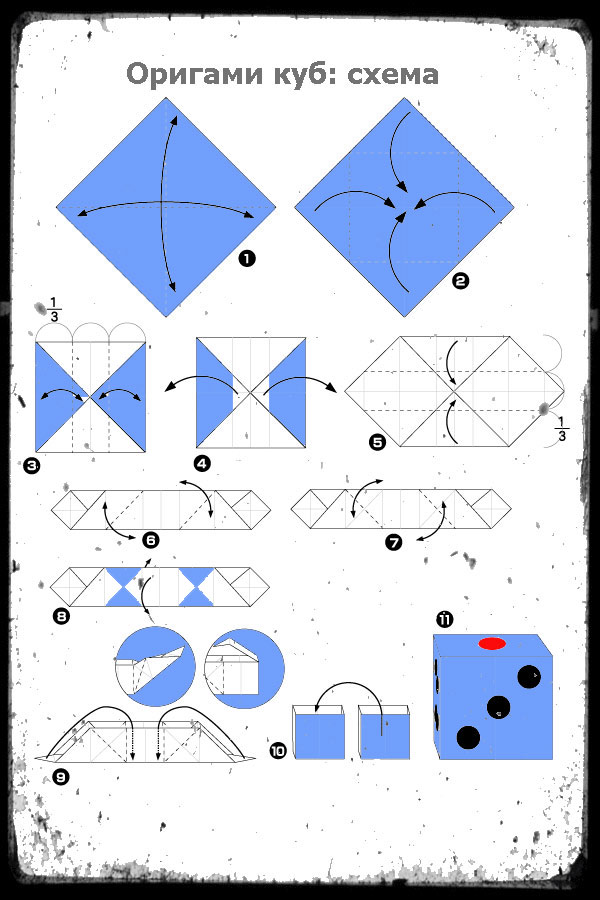
Доступность данных: Код можно найти на GitHub: https://github.com/cistib/origami Данные можно найти на FigShare: https://doi.org/10.6084/m9.figshare.14531421.v1.
Финансирование: Работа была поддержана проектным грантом Исследовательского совета по биотехнологии и биологическим наукам (BBSRC; https://bbsrc.ukri.org/) для TTW, SB и AFF (BB/M01021X/1). Визуализация проводилась в Центре световой микроскопии Вольфсона в Университете Шеффилда при поддержке Исследовательского совета по биотехнологии и биологическим наукам BBSRC ALERT14, присуждаемого TTW и SB за световую микроскопию (BB/M012522/1).AAJ финансировался за счет премии для докторантов от Исследовательского совета по биотехнологии и биологическим наукам BBSRC Партнерство по обучению докторантов White Rose в области механистической биологии (BB / M011151 / 1). AFF поддерживается Кафедрой новых технологий Королевской инженерной академии (CiET1819\19) и сетью MedIAN (EP/N026993/1), финансируемой Исследовательским советом по инженерным и физическим наукам (EPSRC; https://epsrc.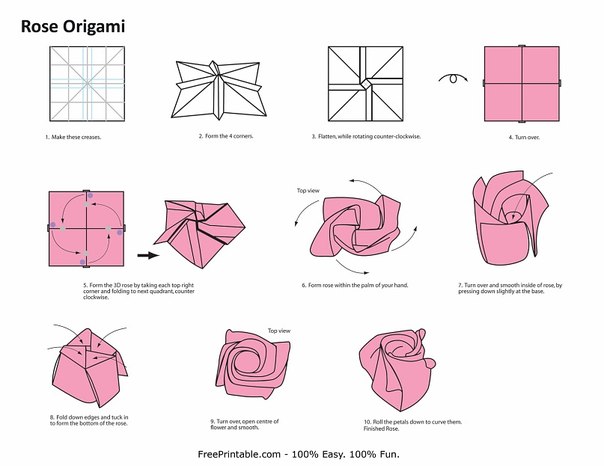 ukri). орг/). Спонсоры не участвовали в разработке исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
ukri). орг/). Спонсоры не участвовали в разработке исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Сложные морфологии различных таксонов и типов тканей возникают в результате деформации эпителиальных пластов [1–3]. У эмбриона многие развивающиеся эпителии образуют сильно изогнутые поверхности. Процессы складывания эпителия управляются поляризованными механическими силами и включают трехмерные изменения формы на клеточном уровне [4,5]. Методы флуоресцентной визуализации позволили проследить такие изменения формы с клеточным разрешением in vivo и в режиме реального времени [6-8].Эти достижения в области визуализации впоследствии привели к разработке инструментов для количественной оценки эпителиальной динамики, особенно изменений формы клеток.
Многие инструменты анализа изображений, измеряющие изменение формы клеток, были ограничены данными двумерной [9–12] или квази-трехмерной флуоресцентной микроскопии [13].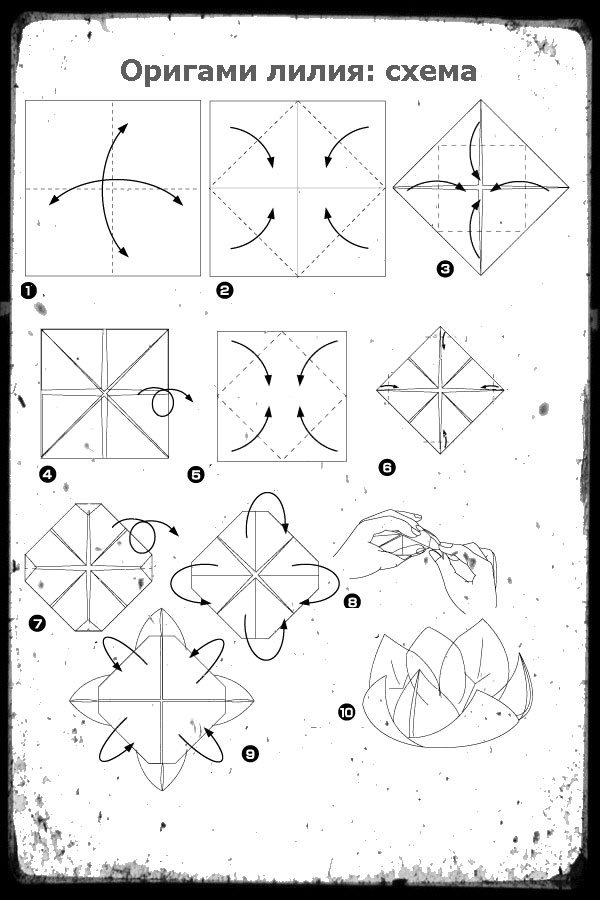 Расширению этих измерений до 3D способствовала разработка основанных на мембранах методов 3D-сегментации, таких как ACME [14], RACE [15], 3DMMS [16], CellProfiler 3.0 [17] и совсем недавно основанных на глубоком обучении. методы [18–21].Некоторые инструменты анализа изображений, такие как CellProfiler 3.0 [17], MorphoGraphX [22] и ShapeMetrics [23], предоставляют конвейеры для вычисления признаков формы клеток, не зависящих от направления. Тем не менее, нахождение положения 3D-сегментированных клеток вдоль биологически значимых осей для количественной оценки характеристик направленной формы все еще остается сложной проблемой, для которой до сих пор не найдено обобщенного решения.
Расширению этих измерений до 3D способствовала разработка основанных на мембранах методов 3D-сегментации, таких как ACME [14], RACE [15], 3DMMS [16], CellProfiler 3.0 [17] и совсем недавно основанных на глубоком обучении. методы [18–21].Некоторые инструменты анализа изображений, такие как CellProfiler 3.0 [17], MorphoGraphX [22] и ShapeMetrics [23], предоставляют конвейеры для вычисления признаков формы клеток, не зависящих от направления. Тем не менее, нахождение положения 3D-сегментированных клеток вдоль биологически значимых осей для количественной оценки характеристик направленной формы все еще остается сложной проблемой, для которой до сих пор не найдено обобщенного решения.
Определение ориентации отдельных клеток относительно известной общей полярности эпителиального слоя имеет решающее значение, поскольку клеточно-поляризованные биомеханические процессы вызывают изменения формы клеток; сужение или расширение может происходить вдоль апикальной [24,25] или базолатеральной [26] клеточной поверхности и может быть обнаружено по любому перекосу в распределении массы внутри клетки вдоль апико-базальной оси симметрии.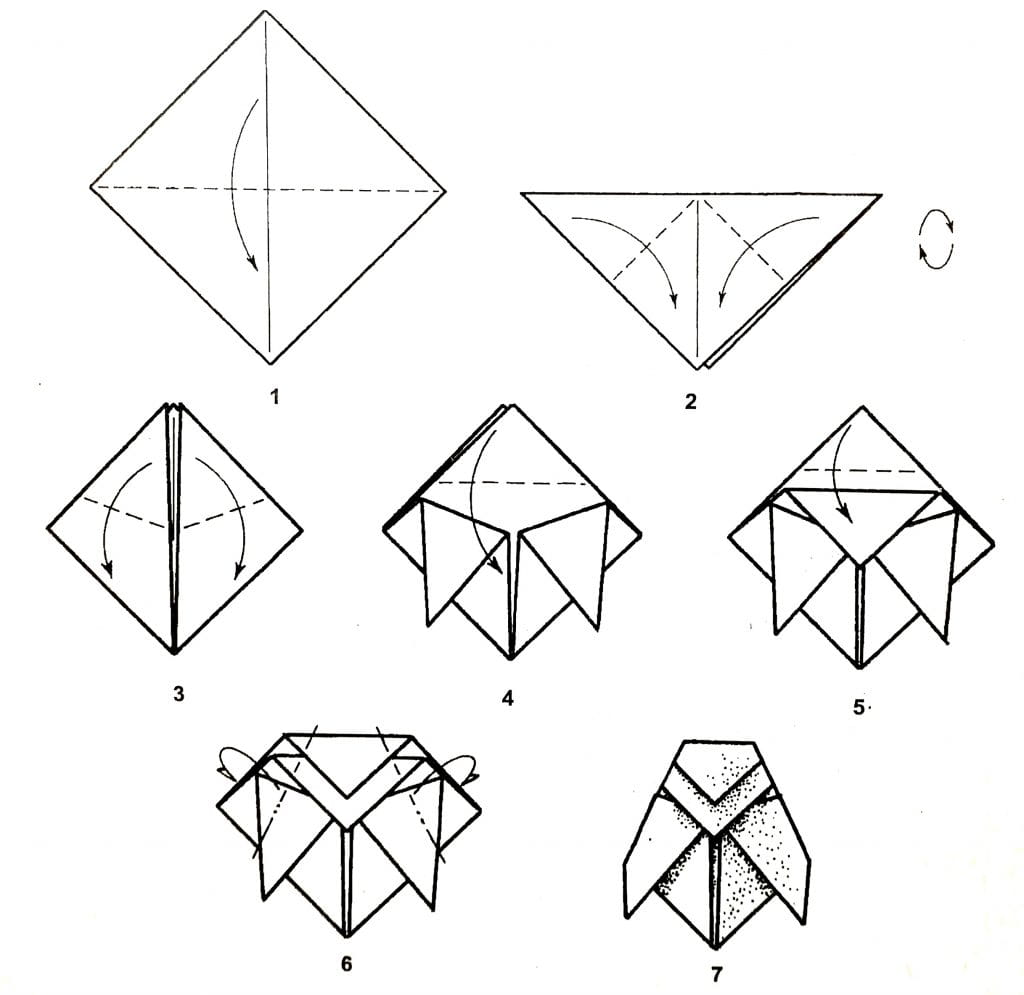 Складывание эпителия может быть инициировано или находиться под влиянием клеточной пролиферации, гибели клеток, ремоделирования цитоскелета или изменений свойств клеточной поверхности [27,28]. Эти механизмы могут привести к изменениям характеристик формы клеток, включая высоту и ширину клеток, объем, площадь поверхности и сферичность.
Складывание эпителия может быть инициировано или находиться под влиянием клеточной пролиферации, гибели клеток, ремоделирования цитоскелета или изменений свойств клеточной поверхности [27,28]. Эти механизмы могут привести к изменениям характеристик формы клеток, включая высоту и ширину клеток, объем, площадь поверхности и сферичность.
Ориентацию или полярность клеток можно определить вдоль плоскости эпителия (плоскостная клеточная полярность) или перпендикулярно плоскости эпителия вдоль апико-базальной оси клетки. Существующие автоматизированные методы определения полярности сегментированных клеток часто полагаются на дополнительные биохимические маркеры полярности [29-31].Включение таких дополнительных маркеров в эксперименты по флуоресцентной визуализации увеличивает время, необходимое для создания каждого изображения, что может привести к фототоксичности, а полученный в результате больший объем данных изображения делает анализ дорогостоящим в вычислительном отношении. Кроме того, производство необходимых животных, несущих несколько трансгенов для живой визуализации, может быть сложным и дорогостоящим. Некоторые методы анализа изображений вычисляют векторы направления для отдельных клеток путем рисования векторов нормалей к полиномиальным функциям, часто эллипсоидам, используемым для оценки поверхности образца — например, целых эмбрионов [15] или бластодерм [32], подвергающихся морфогенезу.Эти методы специфичны для геометрии образца и непригодны для анализа сложных складчатых топологий на продвинутых стадиях морфологического развития. Третий метод использует известные особенности формы клеток для определения ориентации клеток, например, путем применения анализа основных компонентов (PCA) для вычисления апико-базальной оси в столбчатых клетках в EDGE4D [33] и передне-задней оси в зачатках боковой линии рыбок данио. с использованием геометрической морфометрии на основе ориентиров [31] или ориентации клеток вдоль их длинной оси в глазном бокале рыбок данио, как в LongAxis [34].Эти стратегии будут применимы только в том случае, если доминирующая характеристика формы ячейки, например отношение высоты к ширине ячейки, известна и остается неизменной в пространстве и во времени.
Некоторые методы анализа изображений вычисляют векторы направления для отдельных клеток путем рисования векторов нормалей к полиномиальным функциям, часто эллипсоидам, используемым для оценки поверхности образца — например, целых эмбрионов [15] или бластодерм [32], подвергающихся морфогенезу.Эти методы специфичны для геометрии образца и непригодны для анализа сложных складчатых топологий на продвинутых стадиях морфологического развития. Третий метод использует известные особенности формы клеток для определения ориентации клеток, например, путем применения анализа основных компонентов (PCA) для вычисления апико-базальной оси в столбчатых клетках в EDGE4D [33] и передне-задней оси в зачатках боковой линии рыбок данио. с использованием геометрической морфометрии на основе ориентиров [31] или ориентации клеток вдоль их длинной оси в глазном бокале рыбок данио, как в LongAxis [34].Эти стратегии будут применимы только в том случае, если доминирующая характеристика формы ячейки, например отношение высоты к ширине ячейки, известна и остается неизменной в пространстве и во времени.
Мы представляем новый автоматизированный и простой в использовании инструмент Origami для выделения признаков формы с изменяющимся направлением вдоль апико-базальной оси путем реконструкции поверхности эпителия с использованием треугольной сетки (рис. 1). Origami применяется к широкому диапазону геометрии образцов, подвергающихся морфогенезу, и автоматически извлекает векторы направления для отдельных клеток, выровненных по апико-базальной оси эпителиального листа, не требуя дополнительных меток для полярности.Элементы формы с переменным направлением рассчитываются путем вычисления геометрических моментов для объема, заключенного в полигональное представление каждой сегментированной ячейки [35]. Мы демонстрируем универсальность нашего метода, используя данные из ассортимента структур на различных стадиях развития внутри слухового пузырька (развивающегося внутреннего уха) эмбрионов рыбок данио.
Рис. 1. Конвейер анализа изображений Origami.
а. Конфокальная флуоресцентная микрофотография Airyscan (проекция максимальной интенсивности (MIP) 35 z-срезов) развивающегося слухового пузырька рыбки данио на 51. 5 часов после оплодотворения. Красный прямоугольник — передняя проекция; желтый ящик — эндолимфатический мешок; голубой прямоугольник — задняя проекция. ROI расширяются рядом — MIP верхнего ряда и отдельные срезы нижнего ряда. Шкала баров: 20 мкм. Синие стрелки отмечают направление апикобазальной полярности (указывая на апикальную сторону). б. Назначение полярности для сегментированных данных; ROI, окружающая переднюю проекцию, была сегментирована (здесь наложена на MIP) с использованием ACME, центроиды были созданы для каждой сегментированной клетки, и из этих центроидов была создана треугольная поверхностная сетка.Векторы нормалей (синие стрелки) к этой поверхностной сетке представляют апико-базальную ось. в. Характеристики формы клеток рассчитывали относительно заданной апико-базальной оси; здесь выделены три примерных ячейки вместе с 3D-рендерингом, показывающим их положение в передней проекции и соответствующие показатели формы в таблице.
5 часов после оплодотворения. Красный прямоугольник — передняя проекция; желтый ящик — эндолимфатический мешок; голубой прямоугольник — задняя проекция. ROI расширяются рядом — MIP верхнего ряда и отдельные срезы нижнего ряда. Шкала баров: 20 мкм. Синие стрелки отмечают направление апикобазальной полярности (указывая на апикальную сторону). б. Назначение полярности для сегментированных данных; ROI, окружающая переднюю проекцию, была сегментирована (здесь наложена на MIP) с использованием ACME, центроиды были созданы для каждой сегментированной клетки, и из этих центроидов была создана треугольная поверхностная сетка.Векторы нормалей (синие стрелки) к этой поверхностной сетке представляют апико-базальную ось. в. Характеристики формы клеток рассчитывали относительно заданной апико-базальной оси; здесь выделены три примерных ячейки вместе с 3D-рендерингом, показывающим их положение в передней проекции и соответствующие показатели формы в таблице.
https://doi.org/10. 1371/journal.pcbi.1009063.g001
1371/journal.pcbi.1009063.g001
Дизайн и внедрение
Конвейеру Origami предшествует этап сегментации на основе мембраны.Для этого мы использовали программное обеспечение сегментации ACME с открытым исходным кодом [14]. Сегментированные данные подвергаются двум основным операциям в Origami; назначение вектора направления эпителиальной полярности (рис. 1B) и выделение признаков формы (рис. 1C).
Назначение направления полярности отдельным ячейкам
Чтобы вычислить особенности формы клеток, изменяющиеся по направлениям, такие как асимметрия (асимметрия клеточной массы), а также продольное и поперечное распространение, положение сегментированных клеток должно быть найдено в трехмерном пространстве вдоль биологически значимой оси — мы выбрали известную апико-базальную ось. клетки.Складчатый эпителий был реконструирован in silico в виде тонкой «корочки» — сетки с открытой поверхностью, которая триангулирует центроиды сегментированных клеток в трехмерном пространстве с использованием алгоритма корки [36,37] (рис. 1В). Метод Коры вычисляет сетку поверхности из неорганизованных точек — центроидов ячеек в нашем случае, используя диаграмму Вороного центроидов ячеек.
1В). Метод Коры вычисляет сетку поверхности из неорганизованных точек — центроидов ячеек в нашем случае, используя диаграмму Вороного центроидов ячеек.
После этого наш автоматизированный метод исправляет дефекты расчетной сетки поверхности, которые могут привести к ошибкам в результирующих векторах направлений.Сетка уточняется путем удаления дубликатов (в вычисляемых вершинах или треугольных гранях) и любых самопересекающихся треугольных граней. Ребра, не являющиеся многообразиями, то есть ребра, общие для более чем двух треугольных граней, повторно объединяются в сетку многообразия с использованием алгоритма поворота шара [38,39].
Треугольные грани уточненной сетки упорядочены, поэтому, применяя правило правой руки при создании векторов нормалей к поверхностной сетке, все эти векторы указывают на одну и ту же сторону репрезентации сетки эпителиальной поверхности (рис. 1В).На данный момент все еще есть две возможные противоположные ориентации для каждого рассчитанного вектора — обращенные к апикальной или базальной стороне эпителия, с разницей только в знаке. В развивающихся слуховых пузырьках рыбок данио апикальная поверхность эпителия обращена к заполненному жидкостью просвету пузырька [2,8,40]. Мы использовали это предварительное знание, чтобы сообщить ориентацию векторов, установив значение двоичного параметра, определяющего ориентацию, на «внутрь», чтобы они указывали на точку схождения, которая приходится на сторону криволинейной поверхностной сетки, которая соответствует вершине. (люменальная) сторона эпителия каждой клетки.Когда структура складывается несколько раз в противоположных ориентациях, например, в синтетических данных, созданных для этого исследования, «in» устанавливает векторы направления полярности так, чтобы они указывали на глобальную точку схождения, в нашем случае определяемую кривизной всего синтетического эпителия.
В развивающихся слуховых пузырьках рыбок данио апикальная поверхность эпителия обращена к заполненному жидкостью просвету пузырька [2,8,40]. Мы использовали это предварительное знание, чтобы сообщить ориентацию векторов, установив значение двоичного параметра, определяющего ориентацию, на «внутрь», чтобы они указывали на точку схождения, которая приходится на сторону криволинейной поверхностной сетки, которая соответствует вершине. (люменальная) сторона эпителия каждой клетки.Когда структура складывается несколько раз в противоположных ориентациях, например, в синтетических данных, созданных для этого исследования, «in» устанавливает векторы направления полярности так, чтобы они указывали на глобальную точку схождения, в нашем случае определяемую кривизной всего синтетического эпителия.
Недосегментация может привести к отсутствию областей или нежелательным отверстиям в треугольной сетке, что приведет к ошибкам при упорядочении треугольных граней. Наш пайплайн пытается исправить эти дыры, обнаруживая их, а затем перестраивая, где это возможно. Дыры при обнаружении помечаются как предупреждение пользователям о возможных ошибках в выводе. Векторы нормали к реконструированной поверхности представляют собой апико-базальную ось эпителия и генерируются для каждой сегментированной клетки в их центроидном положении (рис. 1B и 1C).
Дыры при обнаружении помечаются как предупреждение пользователям о возможных ошибках в выводе. Векторы нормали к реконструированной поверхности представляют собой апико-базальную ось эпителия и генерируются для каждой сегментированной клетки в их центроидном положении (рис. 1B и 1C).
Вычисление элементов формы с использованием трехмерных геометрических моментов
Форма объекта может быть охарактеризована центральными геометрическими моментами [41]. Геометрические моменты широко используются в задачах распознавания и классификации объектов [42, 43], поскольку они (i) просты для вычисления, (ii) организуют признаки в порядке возрастания детализации и (iii) могут быть расширены до n измерений.Каждый момент определяется интегралом по объекту (в нашем случае по каждой сегментированной ячейке) монома декартовых координат , где i , j , k ≥ 0, с началом координат в центроид.
В нашем конвейере анализа трехмерные геометрические моменты вычислялись с использованием алгоритма, представленного в [35]. Определяющие непрерывные интегралы точно вычисляются в треугольной сетке поверхности, созданной для каждой отдельной сегментированной ячейки, разделенной на сумму:
(1)
где каждый тетраэдр T c определяется треугольником в сетке поверхности и началом координат (центроидом ячейки).Определитель дает ориентированный объем этого тетраэдра,
(2)
Определяющие непрерывные интегралы точно вычисляются в треугольной сетке поверхности, созданной для каждой отдельной сегментированной ячейки, разделенной на сумму:
(1)
где каждый тетраэдр T c определяется треугольником в сетке поверхности и началом координат (центроидом ячейки).Определитель дает ориентированный объем этого тетраэдра,
(2)
Определитель с учетом своего знака позволяет применять алгоритм к фигурам любой сложности и топологии. Интеграл в каждом T c задается замкнутой формулой, включающей только декартовы координаты треугольных вершин.
Геометрические моменты низших порядков имеют простую интуитивную интерпретацию. Нулевой -й -й момент порядка обеспечивает объем объекта, здесь отдельной ячейки.Для центральных моментов моменты первого порядка тривиально равны нулю: . Моменты второго порядка соответствуют разбросу (тензору ковариации) распределения. Таким образом, проекция массы каждой клетки вдоль соответствующего вектора полярности представляет «разброс» как дисперсию массы «продольно» (вдоль апико-базальной оси) и «поперечно» (вдоль эпителиальной плоскости). Это позволило нам определить, были ли клетки более или менее столбчатыми (высокие клетки) или плоскоклеточными (плоские клетки) по форме. Моменты третьего порядка представляют «асимметрию», то есть отклонение от симметрии.В нашем конвейере асимметрия измерялась вдоль вектора направления полярности в апикально-базальном направлении, при этом положительные значения асимметрии указывали на сужение апикальной клетки и/или базальную релаксацию, а отрицательные значения указывали на сужение базальной клетки и/или апикальное расширение. Нулевое значение указывает на отсутствие перекоса. Кроме того, сферичность каждой ячейки рассчитывалась как отношение площади поверхности клетки к площади поверхности сферы того же объема, что и ячейка [44], от 0 для клетки очень неправильной формы до 1 для идеальной сферы. .
Это позволило нам определить, были ли клетки более или менее столбчатыми (высокие клетки) или плоскоклеточными (плоские клетки) по форме. Моменты третьего порядка представляют «асимметрию», то есть отклонение от симметрии.В нашем конвейере асимметрия измерялась вдоль вектора направления полярности в апикально-базальном направлении, при этом положительные значения асимметрии указывали на сужение апикальной клетки и/или базальную релаксацию, а отрицательные значения указывали на сужение базальной клетки и/или апикальное расширение. Нулевое значение указывает на отсутствие перекоса. Кроме того, сферичность каждой ячейки рассчитывалась как отношение площади поверхности клетки к площади поверхности сферы того же объема, что и ячейка [44], от 0 для клетки очень неправильной формы до 1 для идеальной сферы. .
Результаты
Оценка вычисленного вектора направления полярности ячейки
Чтобы оценить вычисленные векторы направления, обозначающие клеточную полярность, мы создали трехмерные синтетические данные, представляющие изогнутые складчатые эпителии с различной степенью кривизны и высотой складчатого пика в двух противоположных ориентациях (текст S1 и рис. 2A). Чтобы отразить реальные условия флуоресцентной визуализации in vivo , эти синтетические данные были искажены тремя инкрементными уровнями гауссовского и пуассоновского шума (текст S1 и рис. 2A).Используя синтетические данные, были оценены два типа ошибок в вычисленных векторах направления полярности: (1) ошибка переключения ориентации, измеренная как процент векторов полярности с противоположной ориентацией (противоположным знаком) истинной полярности (текст S1), и (2) точность направления, измеряемая как средний угол отклонения между векторами полярности, созданными Origami, правильно ориентированными, и истинной полярностью.
2A). Чтобы отразить реальные условия флуоресцентной визуализации in vivo , эти синтетические данные были искажены тремя инкрементными уровнями гауссовского и пуассоновского шума (текст S1 и рис. 2A).Используя синтетические данные, были оценены два типа ошибок в вычисленных векторах направления полярности: (1) ошибка переключения ориентации, измеренная как процент векторов полярности с противоположной ориентацией (противоположным знаком) истинной полярности (текст S1), и (2) точность направления, измеряемая как средний угол отклонения между векторами полярности, созданными Origami, правильно ориентированными, и истинной полярностью.
Рис. 2. Оценка назначения полярности.
а.Поверхностные сетки синтетического эпителия для проверки конвейера анализа Origami. Наряду с этим 3D-рендеринг одного из синтетических эпителиев (вверху) и одного 2D-среза через него (внизу). Каждый том изображения был искажен тремя уровнями шума. б. Взаимосвязь между геометрией поверхности/шумом и качеством сегментации. Столбики погрешностей представляют собой стандартное отклонение. Попарные сравнения Тьюки со значимыми значениями, отмеченными звездочками: оценка игральной кости при радиусе кривизны 200 мкм (1000 пикселей) по сравнению с показателем при 106 мкм (530 пикселей) – p = 0.0004, балл Dice при самом высоком уровне шума по сравнению с самым низким: p = 0,0045. в. Влияние качества сегментации на ошибки при переключении ориентации (слева) и смещении направления в вычисленных векторах полярности (полосы ошибок, выделенные серым цветом, представляют собой стандартное отклонение). Пунктирные линии представляют собой квадратичное и линейное приближение к данным соответственно. д. Плотность вероятности ошибок в направлении полярности в реальных данных флуоресценции эмбрионов рыбок данио. Каждая точка представляет собой процентную ошибку для трехмерного сегментированного объема (n = 27; всего 949 сегментированных ячеек на всех изображениях).Пунктирная линия показывает среднюю ошибку в наборе данных (<4%).
Столбики погрешностей представляют собой стандартное отклонение. Попарные сравнения Тьюки со значимыми значениями, отмеченными звездочками: оценка игральной кости при радиусе кривизны 200 мкм (1000 пикселей) по сравнению с показателем при 106 мкм (530 пикселей) – p = 0.0004, балл Dice при самом высоком уровне шума по сравнению с самым низким: p = 0,0045. в. Влияние качества сегментации на ошибки при переключении ориентации (слева) и смещении направления в вычисленных векторах полярности (полосы ошибок, выделенные серым цветом, представляют собой стандартное отклонение). Пунктирные линии представляют собой квадратичное и линейное приближение к данным соответственно. д. Плотность вероятности ошибок в направлении полярности в реальных данных флуоресценции эмбрионов рыбок данио. Каждая точка представляет собой процентную ошибку для трехмерного сегментированного объема (n = 27; всего 949 сегментированных ячеек на всех изображениях).Пунктирная линия показывает среднюю ошибку в наборе данных (<4%). е. Чувствительность показателей формы клеток к ошибкам ориентации полярности. Точки данных на графиках отображаются тем же цветом, что и соответствующая синтетическая ячейка рядом. парные сравнения Тьюки со значимыми значениями, отмеченными звездочками; Продольный разброс: 1–2 p = 0,039, 1–3 p < 0,0001, 2–3 p < 0,0001; Поперечный разброс: 1–2 p < 0,0001, 1–3 p < 0.0001, 2–3 р < 0,0001; Асимметрия: 1–2 p < 0,0001, 1–3 p < 0,0001, 2–3 p < 0,0001.
е. Чувствительность показателей формы клеток к ошибкам ориентации полярности. Точки данных на графиках отображаются тем же цветом, что и соответствующая синтетическая ячейка рядом. парные сравнения Тьюки со значимыми значениями, отмеченными звездочками; Продольный разброс: 1–2 p = 0,039, 1–3 p < 0,0001, 2–3 p < 0,0001; Поперечный разброс: 1–2 p < 0,0001, 1–3 p < 0.0001, 2–3 р < 0,0001; Асимметрия: 1–2 p < 0,0001, 1–3 p < 0,0001, 2–3 p < 0,0001.
https://doi.org/10.1371/journal.pcbi.1009063.g002
Из двух проанализированных аспектов геометрии поверхности высота изогнутого пика (в двух противоположных направлениях) не вносила значительного вклада в ошибки переключения ориентации (линейная регрессия; p = 0,86, R 2 = -0,04). Однако больший радиус кривизны эпителия (более плоский эпителиальный лист) коррелировал с ошибками переключения ориентации, хотя и с небольшим эффектом 0. 08% увеличение на каждый 1 мкм (5 пикселей) увеличение радиуса кривизны (линейная регрессия; p = 0,042, R 2 = 0,12, эффект) и более низкое качество сегментации на выходе из ACME (линейная регрессия p < 0,001, R 2 = 0,46; рис. 2B), рассчитанный как оценка Dice. Это означало снижение оценки Dice на 0,2% на каждый 1 мкм (5 пикселей) увеличения радиуса кривизны. Эта корреляция может быть связана со сниженной способностью ACME сегментировать плоские плоскоклеточные клетки в эпителии, ориентированном в основном вдоль латеральной ( xy ) плоскости в данных с анизотропным воксельным разрешением (здесь смоделировано с использованием анизотропной функции рассеяния точки (PSF)) .Мы обнаружили корреляцию между шумом, примененным к синтетическим изображениям, и ошибками при переключении ориентации обеих полярностей (ANOVA: p ≈ 0,001; контрасты Тьюки показали увеличение ошибок на 11,3% при самом высоком уровне шума по сравнению с самым низким уровнем шума: p ).
08% увеличение на каждый 1 мкм (5 пикселей) увеличение радиуса кривизны (линейная регрессия; p = 0,042, R 2 = 0,12, эффект) и более низкое качество сегментации на выходе из ACME (линейная регрессия p < 0,001, R 2 = 0,46; рис. 2B), рассчитанный как оценка Dice. Это означало снижение оценки Dice на 0,2% на каждый 1 мкм (5 пикселей) увеличения радиуса кривизны. Эта корреляция может быть связана со сниженной способностью ACME сегментировать плоские плоскоклеточные клетки в эпителии, ориентированном в основном вдоль латеральной ( xy ) плоскости в данных с анизотропным воксельным разрешением (здесь смоделировано с использованием анизотропной функции рассеяния точки (PSF)) .Мы обнаружили корреляцию между шумом, примененным к синтетическим изображениям, и ошибками при переключении ориентации обеих полярностей (ANOVA: p ≈ 0,001; контрасты Тьюки показали увеличение ошибок на 11,3% при самом высоком уровне шума по сравнению с самым низким уровнем шума: p ).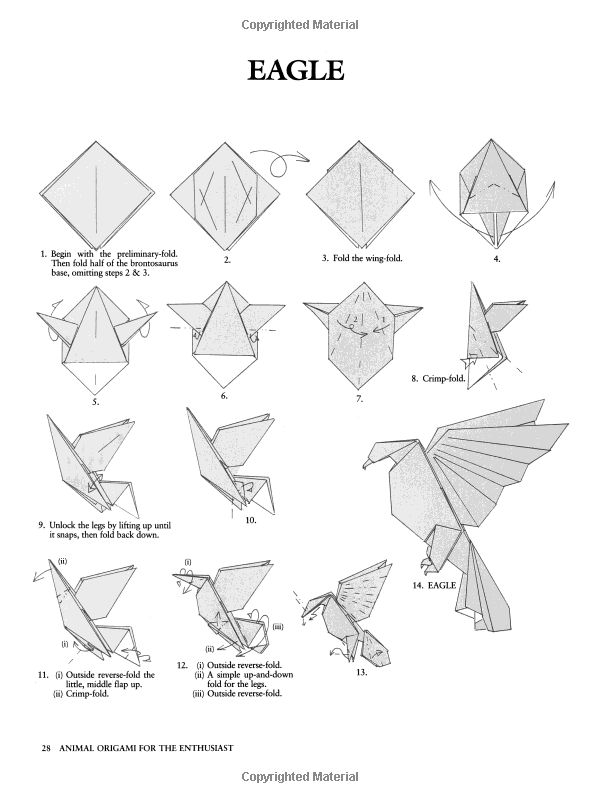 = 0,0039) и результат сегментации (ANOVA: p <0,01; контрасты Тьюки показали снижение оценки Дайса на 16,3% при самом высоком уровне шума по сравнению с самым низким применяемым уровнем шума: p = 0,0045). Качество сегментации, в свою очередь, влияло на переключение полярности с ошибками менее 1.5% при баллах Дайса выше 0,8, но увеличивается при дальнейшем снижении баллов Дайса (полиномиальная регрессия; первый порядок: p <0,001, размер эффекта = -28,78; второй порядок: p <0,01, размер эффекта = 16,26 ; рис. 2С). Сравнение многих доступных алгоритмов сегментации при проверке флуоресцентными изображениями несвернутых структур, таких как зародыши нематод на ранней стадии [16] или корни растений [18], показало, что баллы Дайса превышают 80%, что свидетельствует о хорошей производительности в реальных экспериментальных условиях. условия.
= 0,0039) и результат сегментации (ANOVA: p <0,01; контрасты Тьюки показали снижение оценки Дайса на 16,3% при самом высоком уровне шума по сравнению с самым низким применяемым уровнем шума: p = 0,0045). Качество сегментации, в свою очередь, влияло на переключение полярности с ошибками менее 1.5% при баллах Дайса выше 0,8, но увеличивается при дальнейшем снижении баллов Дайса (полиномиальная регрессия; первый порядок: p <0,001, размер эффекта = -28,78; второй порядок: p <0,01, размер эффекта = 16,26 ; рис. 2С). Сравнение многих доступных алгоритмов сегментации при проверке флуоресцентными изображениями несвернутых структур, таких как зародыши нематод на ранней стадии [16] или корни растений [18], показало, что баллы Дайса превышают 80%, что свидетельствует о хорошей производительности в реальных экспериментальных условиях. условия.
Количественная точность направления была оценена в синтетических данных, для которых, в отличие от данных реальных флуоресцентных изображений, можно было получить надежную базовую истину из известных функций подстилающей поверхности.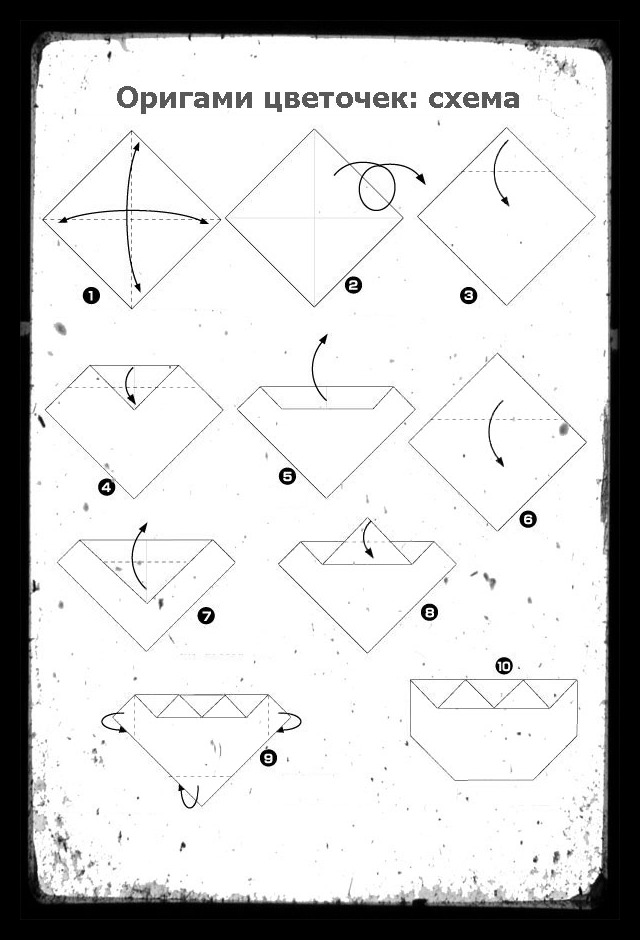 По сравнению с наземными данными полярности, общее смещение 10,6° ± 15,5° (среднее ± стандартное) было измерено из всего нашего синтетического набора данных. Так же, как и для ошибки переключения ориентации полярности, высота свернутого пика не влияла на точность направления полярности (линейная регрессия; p = 0.39, R 2 = -0,01), но наблюдался небольшой эффект кривизны эпителия с дополнительным смещением на 0,06° на каждый 1 мкм (5 пикселей) увеличения радиуса кривизны эпителия (линейная регрессия; р = 0,005, R 2 = 0,24). При самом высоком уровне шума ошибки ориентации полярности имели смещение на 6,6° больше, чем при самом низком уровне шума (контрасты Тьюки; p = 0,003). Был также отрицательный линейный эффект качества сегментации с 2.Смещение на 9° прогнозировалось для каждых 10% снижения оценки Dice (линейная регрессия; p < 0,0001, R 2 = 0,53; рис. 2C).
По сравнению с наземными данными полярности, общее смещение 10,6° ± 15,5° (среднее ± стандартное) было измерено из всего нашего синтетического набора данных. Так же, как и для ошибки переключения ориентации полярности, высота свернутого пика не влияла на точность направления полярности (линейная регрессия; p = 0.39, R 2 = -0,01), но наблюдался небольшой эффект кривизны эпителия с дополнительным смещением на 0,06° на каждый 1 мкм (5 пикселей) увеличения радиуса кривизны эпителия (линейная регрессия; р = 0,005, R 2 = 0,24). При самом высоком уровне шума ошибки ориентации полярности имели смещение на 6,6° больше, чем при самом низком уровне шума (контрасты Тьюки; p = 0,003). Был также отрицательный линейный эффект качества сегментации с 2.Смещение на 9° прогнозировалось для каждых 10% снижения оценки Dice (линейная регрессия; p < 0,0001, R 2 = 0,53; рис. 2C).
Далее мы проверили влияние таких ошибок в точности направления на метрики формы, изменяющиеся в направлении, вычисленные путем применения направленного шума — со средним значением, равным измеренной средней ошибке выше — к векторам полярности трех ячеек в качестве примера, демонстрирующих экстремальные особенности формы из синтетического набор данных и вычисленные метрики формы в зависимости от направления для каждого нового смещенного вектора полярности ( n = 50; рис. 2E).Результирующие вычисленные метрики формы по-прежнему могут успешно различать три ячейки, показывая, что ошибки точности направления (исключая ошибки изменения ориентации) не должны неблагоприятно влиять на вычисленные метрики формы. С другой стороны, ошибки переворота ориентации будут влиять на метрики формы, но, как показано выше, эти ошибки прогнозируются небольшими для хорошо сегментированного объема изображения и могут быть легко идентифицированы при визуальном осмотре и при необходимости исправлены с помощью конвейера Origami.
2E).Результирующие вычисленные метрики формы по-прежнему могут успешно различать три ячейки, показывая, что ошибки точности направления (исключая ошибки изменения ориентации) не должны неблагоприятно влиять на вычисленные метрики формы. С другой стороны, ошибки переворота ориентации будут влиять на метрики формы, но, как показано выше, эти ошибки прогнозируются небольшими для хорошо сегментированного объема изображения и могут быть легко идентифицированы при визуальном осмотре и при необходимости исправлены с помощью конвейера Origami.
Кроме того, ошибки переключения ориентации были количественно оценены по данным реальной флуоресцентной микроскопии светового листа структур в развивающихся отических пузырьках рыбок данио (рис. 1 и 3). Для этого клетки, которым была присвоена неправильная ориентация вдоль апико-базальной оси, то есть обращенные к базальной поверхности, а не к апикальной поверхности, были идентифицированы с помощью визуальной оценки в конвейере Оригами, показывая ошибки в 3,65% из n = 949 клеток.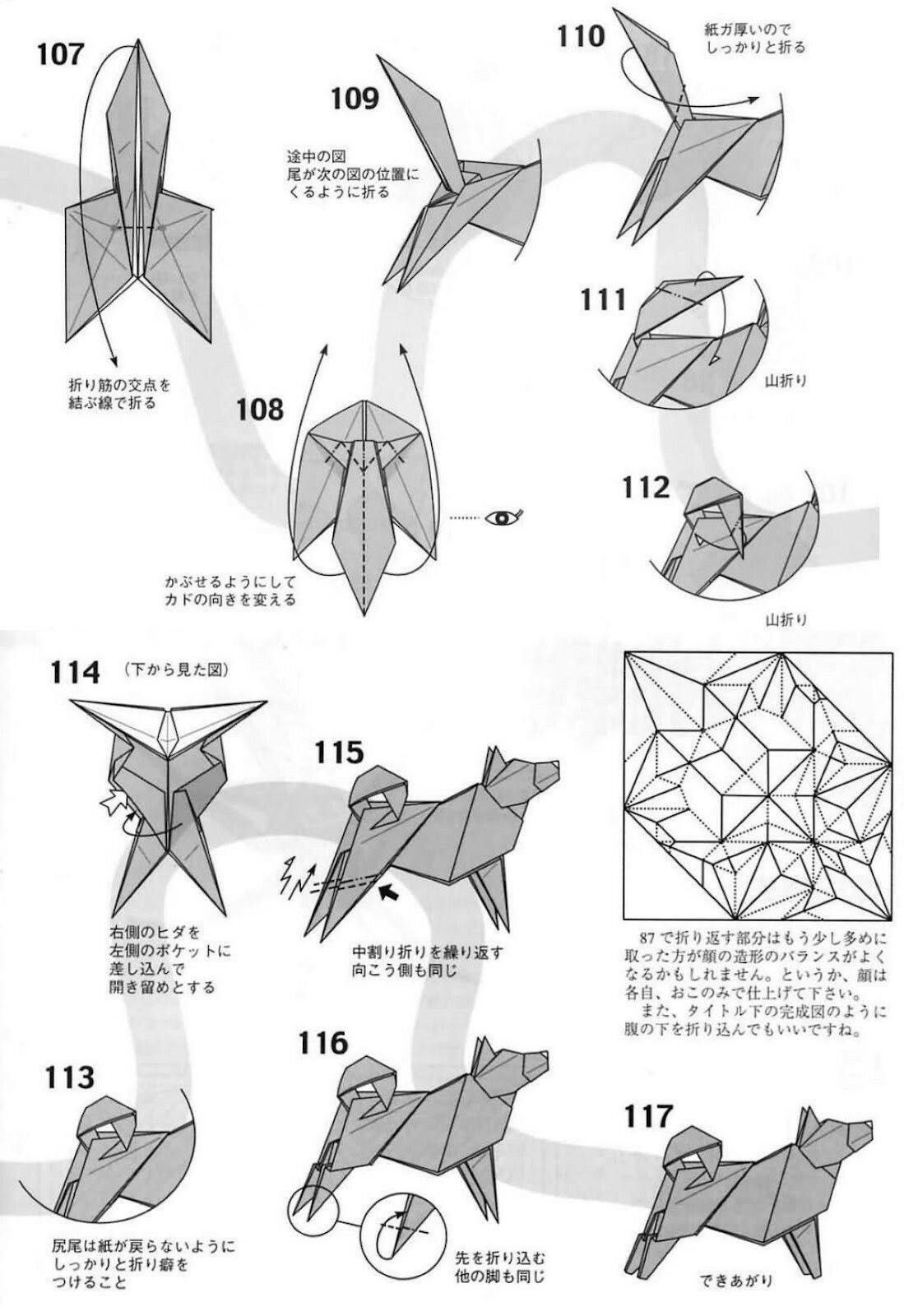 проанализированы (рис. 2D).
проанализированы (рис. 2D).
Рис. 3. Сравнение динамики формы развивающихся структур внутреннего уха рыбок данио.
Строки представляют каждый анализируемый момент времени. Данные, выделенные синим цветом, представляют собой клетки из AP, зеленые представляют собой клетки из PP, а пурпурный цвет представляют собой ES. а. Двойные диаграммы линейного дискриминационного анализа (LDA) иллюстрируют многомерную кластеризацию данных: данные из AP и PP показывают значительное перекрытие, указывающее на схожие сигнатуры формы, в то время как данные из ES показывают меньшее совпадение с первым. б. Схематические иллюстрации сигнатур формы клеток в проанализированные моменты времени, показывающие клетки в ES, имеющие перекос в направлении, противоположном таковым в проекциях, и имеющие менее округлые формы.Стрелки указывают на апико-базальную полярность. в. Графики, показывающие различия в асимметрии и сферичности между структурами в проанализированные моменты времени. Желтые точки с линиями ошибок представляют собой среднее значение и стандартное отклонение для данных. p значения для парных сравнений взяты из таблицы 1.
p значения для парных сравнений взяты из таблицы 1.
https://doi.org/10.1371/journal.pcbi.1009063.g003
Доказательство принципа: понимание динамики формы клеток во время эпителиального морфогенеза во внутреннем ухе рыбок данио
Для дальнейшего подтверждения нашего метода мы использовали Origami для характеристики динамики формы клеток, участвующих в формировании различных структур в слуховом пузырьке эмбриона рыбки данио (рис. 1 и 3).Мы проанализировали данные флуоресцентного изображения светового листа из передней эпителиальной проекции (AP) для развивающейся системы полукружных каналов вместе с эндолимфатическим мешком (ES) в трех временных точках развития: 42,5 часа после оплодотворения (hpf) (момент времени 1) , 44,5 hpf (момент времени 2) и 50,5 hpf (момент времени 3), используя трех разных рыб для каждого момента времени. Мы также проанализировали заднюю эпителиальную проекцию (PP), структуру, аналогичную AP, но которая развивается позже [40], в моменты времени, эквивалентные таковым для AP (46.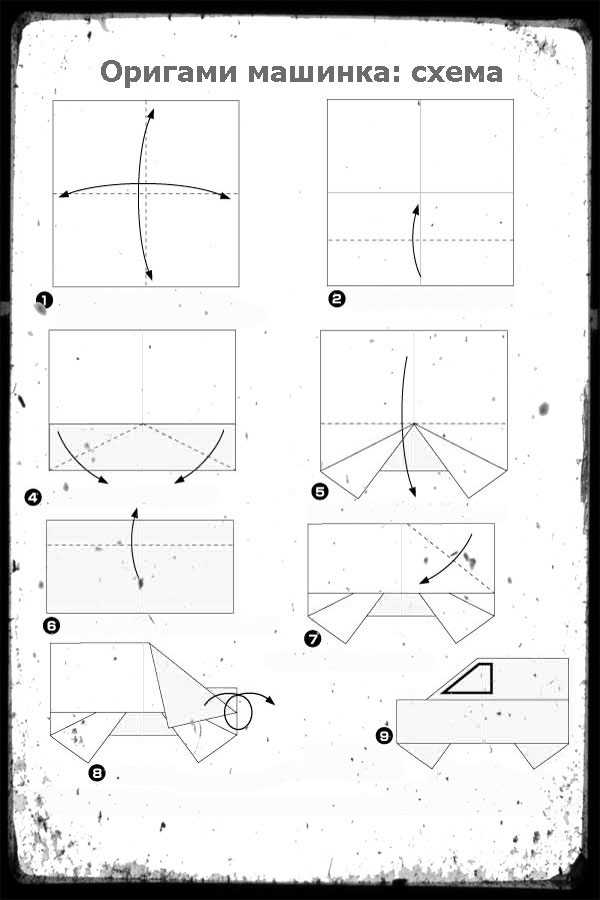 5 л.с., 50,5 л.с. и 60,5 л.с.). AP и PP представляют собой пальцевидные проекции эпителия, которые перемещаются в просвет везикулы, при этом апикальная сторона клетки находится снаружи изогнутой проекционной поверхности [40]. Напротив, ES формируется как инвагинация из дорсального отического эпителия, при этом суженные апикальные поверхности клеток выстилают узкий просвет результирующего короткого протока [8,45,46]. Поскольку ES формируется за счет деформации эпителиального листка с полярностью, противоположной полярности эпителиальных выступов, мы ожидаем, что клетки в ES и выступах будут демонстрировать значительные различия в форме клеток.Наоборот, мы не ожидаем существенных различий в форме клеток между клетками AP и PP, которые формируют эквивалентные структуры в развивающемся ухе.
5 л.с., 50,5 л.с. и 60,5 л.с.). AP и PP представляют собой пальцевидные проекции эпителия, которые перемещаются в просвет везикулы, при этом апикальная сторона клетки находится снаружи изогнутой проекционной поверхности [40]. Напротив, ES формируется как инвагинация из дорсального отического эпителия, при этом суженные апикальные поверхности клеток выстилают узкий просвет результирующего короткого протока [8,45,46]. Поскольку ES формируется за счет деформации эпителиального листка с полярностью, противоположной полярности эпителиальных выступов, мы ожидаем, что клетки в ES и выступах будут демонстрировать значительные различия в форме клеток.Наоборот, мы не ожидаем существенных различий в форме клеток между клетками AP и PP, которые формируют эквивалентные структуры в развивающемся ухе.
Для каждой структуры были вычислены следующие атрибуты формы на уровне одной ячейки: площадь поверхности, сферичность, продольное распространение, поперечное распространение и асимметрия. Поскольку объем и площадь поверхности показывают высокую коллинеарность в наших данных (коэффициент корреляции Пирсона = 0,98, 95% доверительные интервалы = [0,977, 0,984]), объем клеток был исключен из дальнейшего анализа. Хотя изображения включали клетки нескладывающегося эпителия вокруг интересующих развивающихся структур, анализировались только клетки складчатого эпителия. Многофакторный анализ (пакет {MANOVA.RM} [47] в R [v 4.0.0]) зависимости атрибутов формы клеток от эпителиальной структуры, из которой они получены, в разные моменты времени показал значительную разницу между тремя структурами. в первые два периода развития (статистика типа Вальда; p = 0,035 (повторная выборка p = 0.001) в момент времени 1 и p < 0,001 (после повторной выборки p < 0,001) в момент времени 2), но не в последний проанализированный момент времени ( p = 0,706 ( после повторной выборки p = 0,038)) для всех атрибуты формы. Апостериорные контрасты Тьюки показали, что клетки в эндолимфатическом мешке демонстрировали значительно отличающуюся динамику формы от клеток в обеих проекциях (ES-AP p = 0,006, PP-ES p = 0,012 в момент времени 1; ES-AP р = 0.
Хотя изображения включали клетки нескладывающегося эпителия вокруг интересующих развивающихся структур, анализировались только клетки складчатого эпителия. Многофакторный анализ (пакет {MANOVA.RM} [47] в R [v 4.0.0]) зависимости атрибутов формы клеток от эпителиальной структуры, из которой они получены, в разные моменты времени показал значительную разницу между тремя структурами. в первые два периода развития (статистика типа Вальда; p = 0,035 (повторная выборка p = 0.001) в момент времени 1 и p < 0,001 (после повторной выборки p < 0,001) в момент времени 2), но не в последний проанализированный момент времени ( p = 0,706 ( после повторной выборки p = 0,038)) для всех атрибуты формы. Апостериорные контрасты Тьюки показали, что клетки в эндолимфатическом мешке демонстрировали значительно отличающуюся динамику формы от клеток в обеих проекциях (ES-AP p = 0,006, PP-ES p = 0,012 в момент времени 1; ES-AP р = 0. 0002, PP — ES p = 0,0002 в момент времени 2, но ES — AP p = 0,192, PP — ES p = 0,116 в момент времени 3). Не было существенной разницы в сигнатуре формы клеток между клетками в передней и задней проекциях (контрасты Тьюки; PP — AP p = 0,997 в момент времени 1; PP — AP p = 0,999 в момент времени 2 и PP — 0,999). AP p = 0,896 в момент времени 3). Эти результаты показывают, что включенные признаки формы клеток были более похожими, чем разными для клеток из структур в третий проанализированный момент времени.
0002, PP — ES p = 0,0002 в момент времени 2, но ES — AP p = 0,192, PP — ES p = 0,116 в момент времени 3). Не было существенной разницы в сигнатуре формы клеток между клетками в передней и задней проекциях (контрасты Тьюки; PP — AP p = 0,997 в момент времени 1; PP — AP p = 0,999 в момент времени 2 и PP — 0,999). AP p = 0,896 в момент времени 3). Эти результаты показывают, что включенные признаки формы клеток были более похожими, чем разными для клеток из структур в третий проанализированный момент времени.
Из анализируемых признаков: асимметрия (критерий Крускала-Уоллиса; p = 0,008 во время 1, p = 0,004 во время 2 и p < 0,0001 во время 3), сферичность (критерий Крускала-Уоллиса; p p = 0,004 во время 2) < 0,001 во время 1, p = 0,00012 во время 2 и p < 0,001 во время 3) и площадь поверхности (критерий Крускала-Уоллиса; p < 0,001 во время 1, p < 1 во время 0,00067) 2 и p = 0,018 во время 3) описаны значительные различия в форме клеток во всех трех проанализированных временных точках; клетки эндолимфатического мешка характеризовались положительными значениями асимметрии, меньшими значениями сферичности и большей площадью поверхности по сравнению с клетками в обеих проекциях, которые показывают отрицательные значения асимметрии (табл.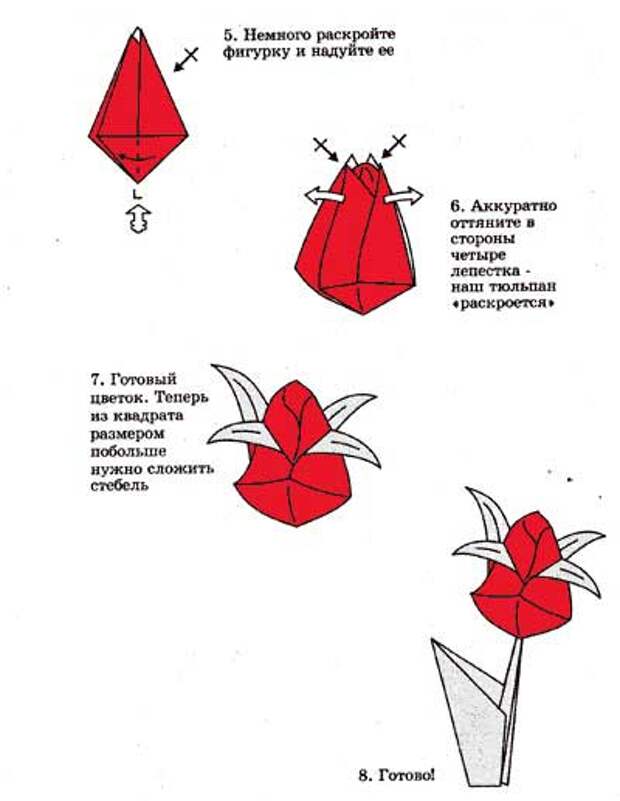 1 и рис. 3).
1 и рис. 3).
Различия в площади поверхности, вероятно, связаны с различиями в сферичности клеток в трех структурах, но не в размерах, поскольку поперечное и продольное распространение не выявило существенных различий.
Доступность и будущие направления
Origami можно бесплатно загрузить с https://github.com/cistib/origami. Он реализован в MATLAB (совместимость с версией 2018b и выше) и включает дополнительные инструменты для визуализации метрик формы клеток из сложных складчатых эпителиев на уровне одной клетки.Инструкции по установке и использованию прилагаются к программному обеспечению.
Наше программное обеспечение может принимать предварительно сегментированные данные, что делает его совместимым с алгоритмами сегментации по выбору пользователя, потенциально позволяя анализировать данные, полученные с использованием других методов трехмерной визуализации, таких как томография. Сегментированные данные должны точно отражать форму клетки, поэтому выбор метода визуализации, который может точно определить трехмерную форму клетки наряду с сегментацией на основе мембраны или цитоплазмы, имеет решающее значение.
Мы использовали априорные знания об организации слухового эпителия, чтобы сообщить ориентацию апико-базальной оси эпителиального листка лицом к просвету слухового пузырька [2,8,40]. Важно знать организацию апико-базальной оси клеток внутри любой новой структуры, изучаемой для применения Оригами, где параметр, определяющий ориентацию, может быть установлен таким образом, чтобы всегда быть «внутри» или «наружу» в зависимости от того, полярность или нет. вектор направления должен указывать на внутреннюю или внешнюю поверхность изогнутой конструкции.Мы также предположили, что отдельные клетки не нарушают эту организацию, так как это невозможно обнаружить без дополнительных меток, специфичных для полярности. В таком случае векторы полярности из нашего анализа могут быть дополнены информацией из маркировки, специфичной для полярности, для отслеживания такого поведения. Кроме того, для вычисления признаков формы вдоль альтернативной оси полярности конвейер может принимать предварительно назначенную полярность в виде списка векторов для конкретной ячейки.
Мы ожидаем, что Origami будет применяться для изучения широкого спектра морфогенетических процессов и внесет свой вклад в наше понимание лежащих в их основе биомеханических процессов.
Быстрое прототипирование произвольного 2D- и 3D-каркаса ДНК-оригами | Исследование нуклеиновых кислот
Аннотация
Сборки Wireframe DNA origami теперь можно программировать автоматически сверху вниз с использованием простой целевой геометрии каркаса или сеток в 2D и 3D, используя либо жесткую связку из шести спиралей (6HB), либо более подходящую связку из двух спиралей (DX ) края. Хотя эти сборки имеют множество применений в производстве наноразмерных материалов из-за их наноразмерной пространственной адресации и высокой степени настройки, пока не существует простого в использовании программного обеспечения с графическим пользовательским интерфейсом для развертывания этих алгоритмических подходов в одном автономном интерфейсе. Кроме того, последовательное проектирование сверху вниз 3D-объектов на основе DX, ранее поддерживаемое DAEDALUS, было ограничено дискретными длинами ребер и одинаковыми углами вершин, что ограничивало объем объектов, которые можно проектировать. Здесь мы представляем пакет программного обеспечения с открытым исходным кодом ATHENA с графическим пользовательским интерфейсом, который автоматически отображает маршрутизацию одноцепочечных каркасов ДНК и последовательности основных цепей для любого целевого каркаса ДНК-оригами с использованием ребер DX или 6HB, включая неправильные, асимметричные полиэдры на основе DX с различные длины ребер и вершин, продемонстрированные экспериментально, что значительно расширяет набор возможных сборок на основе трехмерной ДНК, которые можно спроектировать.ATHENA также позволяет осуществлять внешнее редактирование последовательностей с использованием caDNAno, что демонстрируется с использованием асимметричного наноразмерного позиционирования наночастиц золота, а также предоставление моделей атомного уровня для молекулярной динамики, грубозернистой динамики с oxDNA и других подходов к моделированию вычислительной химии.
Кроме того, последовательное проектирование сверху вниз 3D-объектов на основе DX, ранее поддерживаемое DAEDALUS, было ограничено дискретными длинами ребер и одинаковыми углами вершин, что ограничивало объем объектов, которые можно проектировать. Здесь мы представляем пакет программного обеспечения с открытым исходным кодом ATHENA с графическим пользовательским интерфейсом, который автоматически отображает маршрутизацию одноцепочечных каркасов ДНК и последовательности основных цепей для любого целевого каркаса ДНК-оригами с использованием ребер DX или 6HB, включая неправильные, асимметричные полиэдры на основе DX с различные длины ребер и вершин, продемонстрированные экспериментально, что значительно расширяет набор возможных сборок на основе трехмерной ДНК, которые можно спроектировать.ATHENA также позволяет осуществлять внешнее редактирование последовательностей с использованием caDNAno, что демонстрируется с использованием асимметричного наноразмерного позиционирования наночастиц золота, а также предоставление моделей атомного уровня для молекулярной динамики, грубозернистой динамики с oxDNA и других подходов к моделированию вычислительной химии.
ВВЕДЕНИЕ
Нанотехнология структурной ДНК была задумана в новаторской работе Неда Симана (1), в которой он постулировал, что синтетическая ДНК может быть использована для программирования синтетических материалов с заданными структурными характеристиками нанометрового масштаба.Использование Симаном синтетических олигонуклеотидов привело к получению протяженных, кристаллоподобных самособирающихся материалов на основе ДНК без контроля их общего размера и протяженности. Более двух десятилетий спустя, основываясь на правилах проектирования Симана, Пол Ротемунд представил концепцию ДНК-оригами (2), в которой используется длинный одноцепочечный ДНК-геном фага M13mp18, который он использовал для создания шаблонов от десятков до сотен более коротких комплементарных синтетических ДНК. Нити ДНК, которые самособирались или «сворачивались» с образованием единого дискретного продукта ДНК с высоким выходом. В то время как M13mp18 по-прежнему является наиболее часто используемым каркасом для этой цели, подход Ротемунда в целом применим к любой длине и последовательности каркаса, которые могут быть получены ферментативно (3,4) или бактериально (5,6). Вскоре после изобретения Ротемунда Дуглас и др. (7) применил подход Ротемунда к самостоятельной сборке 3D-объектов на основе аналогичных правил проектирования, а также выпустил широко используемое программное обеспечение с графическим интерфейсом пользователя (GUI) caDNAno (8), чтобы помочь в ручном проектировании этого класса ДНК-оригами, в котором Дуплексы ДНК расположены на параллельных сотовых или квадратных решетках, также называемых «кирпичиками» оригами.В то время как caDNAno оказался чрезвычайно полезным для ручной разводки каркасов и полуавтоматического конструирования последовательностей комплементарных скоб, используемых для самосборки или складывания фигур-мишеней (7–10), он имеет ограниченную полезность для относительно нового класса каркасных сборок ДНК-оригами.
В то время как M13mp18 по-прежнему является наиболее часто используемым каркасом для этой цели, подход Ротемунда в целом применим к любой длине и последовательности каркаса, которые могут быть получены ферментативно (3,4) или бактериально (5,6). Вскоре после изобретения Ротемунда Дуглас и др. (7) применил подход Ротемунда к самостоятельной сборке 3D-объектов на основе аналогичных правил проектирования, а также выпустил широко используемое программное обеспечение с графическим интерфейсом пользователя (GUI) caDNAno (8), чтобы помочь в ручном проектировании этого класса ДНК-оригами, в котором Дуплексы ДНК расположены на параллельных сотовых или квадратных решетках, также называемых «кирпичиками» оригами.В то время как caDNAno оказался чрезвычайно полезным для ручной разводки каркасов и полуавтоматического конструирования последовательностей комплементарных скоб, используемых для самосборки или складывания фигур-мишеней (7–10), он имеет ограниченную полезность для относительно нового класса каркасных сборок ДНК-оригами. которые отображают сложную 2D- и 3D-геометрию с использованием каркасных «сеток», которые могут состоять из одинарных (11–13), двойных (3, 14–16) или шестиспиральных ребер пучка (17–19).
которые отображают сложную 2D- и 3D-геометрию с использованием каркасных «сеток», которые могут состоять из одинарных (11–13), двойных (3, 14–16) или шестиспиральных ребер пучка (17–19).
Каркас ДНК-оригами с использованием двойного кроссовера (DX) и двухспирального пучка (2HB) впервые был реализован Yan et al. (14) с самосборкой плиток, которая позже была обобщена до 2D и 3D ДНК-оригами Zhang et al. (15). Бенсон и др. (11) позже воспроизвел полиэдрическую геометрию в ДНК полуавтоматически, используя одиночные дуплексы, а в 2016 году Veneziano et al. (3) продемонстрировал, что геометрия 3D-каркаса, основанная исключительно на ребрах DX, может быть спроектирована полностью автоматически на основе целевой геометрии с использованием DAEDALUS. Однако DAEDALUS был ограничен объектами, состоящими из ребер, длина которых кратна 10.5 п.н. и одинаковые углы при вершине. В 2019 г. Jun et al. продемонстрировал полностью автоматическую процедуру проектирования сложных 2D-каркасов ДНК-оригами без каких-либо ограничений по длине ребра или геометрической симметрии на основе ребер DX, названную PERDIX (16).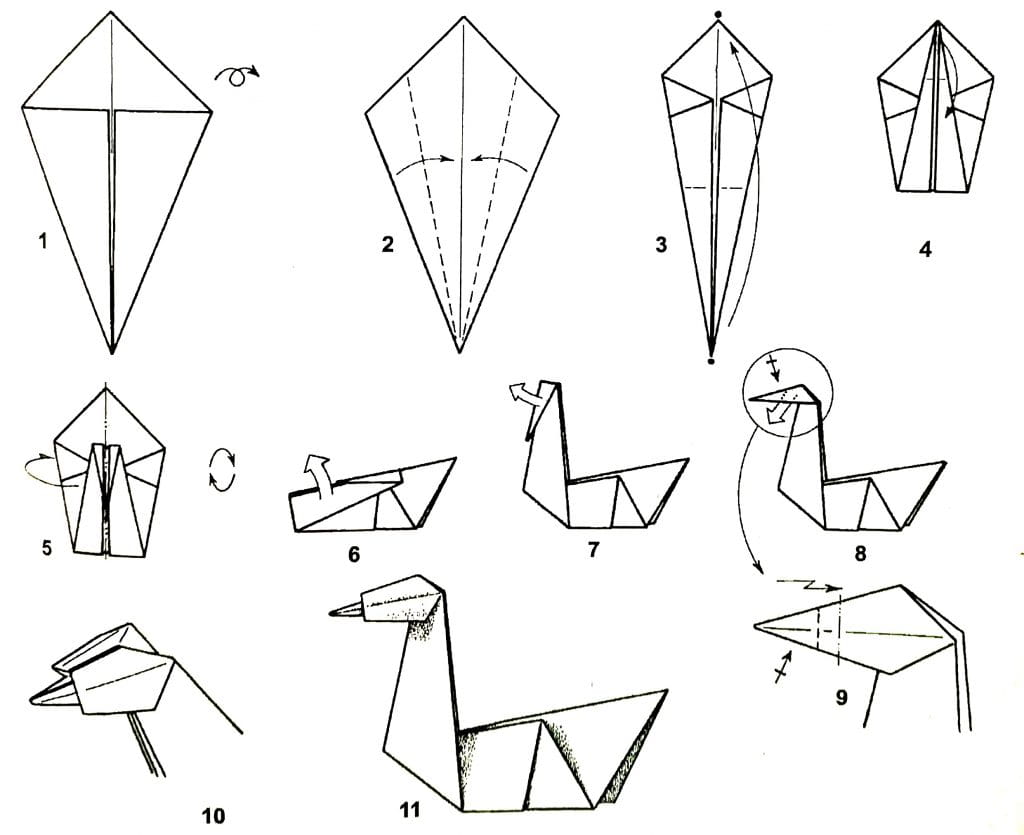 Вскоре после этого они применили аналогичный принцип проектирования для создания краевых 3D-сборок с шестью спиралями (6HB) (TALOS) (17) и 2D-сборок (METIS) (18), где 2D- и 3D-сборки на основе 6HB показали значительно улучшенные характеристики. механическая жесткость по сравнению с аналогами с кромкой DX.PERDIX, METIS и TALOS также позволили визуализировать значительно более широкие классы асимметричных и сложных геометрий за счет введения непрерывных ребер произвольной длины и вершин, подчеркнув возможности каркасного ДНК-оригами для создания сложных наноразмерных материалов с помощью автоматических процедур проектирования. например, в виде наночастиц субъединичной вакцины (20). Однако на сегодняшний день DAEDALUS по-прежнему не может отображать последовательности ДНК для совершенно произвольных трехмерных многогранников, состоящих из недискретных длин ребер и произвольных углов вершин.Кроме того, никакое интегрированное программное обеспечение с графическим интерфейсом, подобное caDNAno (8), не позволяет легко проектировать последовательности ДНК, необходимые для изготовления любого каркасного объекта оригами на основе DAEDALUS, PERDIX, TALOS или METIS.
Вскоре после этого они применили аналогичный принцип проектирования для создания краевых 3D-сборок с шестью спиралями (6HB) (TALOS) (17) и 2D-сборок (METIS) (18), где 2D- и 3D-сборки на основе 6HB показали значительно улучшенные характеристики. механическая жесткость по сравнению с аналогами с кромкой DX.PERDIX, METIS и TALOS также позволили визуализировать значительно более широкие классы асимметричных и сложных геометрий за счет введения непрерывных ребер произвольной длины и вершин, подчеркнув возможности каркасного ДНК-оригами для создания сложных наноразмерных материалов с помощью автоматических процедур проектирования. например, в виде наночастиц субъединичной вакцины (20). Однако на сегодняшний день DAEDALUS по-прежнему не может отображать последовательности ДНК для совершенно произвольных трехмерных многогранников, состоящих из недискретных длин ребер и произвольных углов вершин.Кроме того, никакое интегрированное программное обеспечение с графическим интерфейсом, подобное caDNAno (8), не позволяет легко проектировать последовательности ДНК, необходимые для изготовления любого каркасного объекта оригами на основе DAEDALUS, PERDIX, TALOS или METIS. Adenita (21) — это пакет программного обеспечения с графическим интерфейсом пользователя, который не работает с каркасными сборками 2D или 6HB и опирается на устаревшую версию DAEDALUS, которая ограничена объектами с дискретными длинами ребер и правильными углами вершин.
Adenita (21) — это пакет программного обеспечения с графическим интерфейсом пользователя, который не работает с каркасными сборками 2D или 6HB и опирается на устаревшую версию DAEDALUS, которая ограничена объектами с дискретными длинами ребер и правильными углами вершин.
Чтобы обеспечить полностью автоматизированное проектирование последовательности любой каркасной 2D- или 3D-оригами сборки ДНК с DX- или 6HB-ребрами, с произвольными длинами ребер и углами вершин, здесь мы представляем ATHENA.ATHENA состоит из графического пользовательского интерфейса, который интегрирует эти возможности, а также предлагает фундаментальные алгоритмические усовершенствования, позволяющие использовать произвольные длины и углы ребер для точного проектирования произвольных каркасных объектов, включая асимметричные и неправильные геометрические формы с использованием либо равномерно 6HB (17,18), либо ребер на основе DX ( 3,16). В дополнение к полностью автоматизированному дизайну последовательностей, ATHENA создает выходные файлы, включая структуры всех атомов в формате банка данных белков (PDB) (22) для молекулярной визуализации с использованием таких инструментов, как Visual Molecular Dynamics (VMD) (23) или UCSF Chimera (24). , моделирование молекулярной динамики всех атомов или крупнозернистое моделирование с использованием таких инструментов, как oxDNA (25,26) или mrDNA (27), а также файлы caDNAno для редактирования или модификации конструкций последовательностей для функционализации ДНК-оригами или других целей (28) , а также полные файлы последовательностей для упорядочения цепей основных олигонуклеотидов, необходимых для изготовления с помощью самосборки в одном горшке.Мы проверяем конструкции последовательности для пятиугольных объектов 6HB с использованием атомно-силовой микроскопии (АСМ), просвечивающей электронной микроскопии (ПЭМ) и крупнозернистого моделирования oxDNA, а также для асимметричного октаэдра на основе DX с использованием криоэлектронной микроскопии (крио-ЭМ). Мы дополнительно иллюстрируем использование ATHENA для асимметричного двумерного размещения наночастиц золота (AuNP) с разрешением в нанометровом масштабе.
, моделирование молекулярной динамики всех атомов или крупнозернистое моделирование с использованием таких инструментов, как oxDNA (25,26) или mrDNA (27), а также файлы caDNAno для редактирования или модификации конструкций последовательностей для функционализации ДНК-оригами или других целей (28) , а также полные файлы последовательностей для упорядочения цепей основных олигонуклеотидов, необходимых для изготовления с помощью самосборки в одном горшке.Мы проверяем конструкции последовательности для пятиугольных объектов 6HB с использованием атомно-силовой микроскопии (АСМ), просвечивающей электронной микроскопии (ПЭМ) и крупнозернистого моделирования oxDNA, а также для асимметричного октаэдра на основе DX с использованием криоэлектронной микроскопии (крио-ЭМ). Мы дополнительно иллюстрируем использование ATHENA для асимметричного двумерного размещения наночастиц золота (AuNP) с разрешением в нанометровом масштабе.
МАТЕРИАЛЫ И МЕТОДЫ
Реализация графического интерфейса
ATHENA — это программное приложение с открытым исходным кодом (GNU GPLv3) с графическим интерфейсом пользователя (https://github. com/lcbb/athena), который выполняет полностью автоматизированный дизайн последовательности 2D- и 3D-каркасных ДНК-оригами объектов на основе ребер на основе DX и 6HB и любого интересующего каркаса одноцепочечной ДНК. ATHENA была реализована на Python с использованием библиотек Qt5. Пакеты внутреннего программного обеспечения PERDIX, DAEDALUS, METIS и TALOS встроены в виде двоичных файлов.
com/lcbb/athena), который выполняет полностью автоматизированный дизайн последовательности 2D- и 3D-каркасных ДНК-оригами объектов на основе ребер на основе DX и 6HB и любого интересующего каркаса одноцепочечной ДНК. ATHENA была реализована на Python с использованием библиотек Qt5. Пакеты внутреннего программного обеспечения PERDIX, DAEDALUS, METIS и TALOS встроены в виде двоичных файлов.
Произвольные длины ребер и вершины в DAEDALUS
Автоматический дизайн последовательности на основе DX для любой целевой трехмерной многогранной геометрии с использованием непрерывных длин ребер и произвольных вершин обеспечивается алгоритмическими достижениями в DAEDALUS.Конструкции с непрерывными ребрами позволяют визуализировать объекты на основе ДНК с использованием непрерывных, произвольных длин ребер и углов вершин, при этом один дуплекс заполняет пробел в каждой вершине. Непарные каркасные нуклеотиды используются для охвата расстояния между 3′- и 5′-концами между входящим и исходящим краями, которые в противном случае были бы смещены из-за нативного поворота ДНК B-формы. Вкратце, алгоритм может обрабатывать произвольные длины ребер и углы вершин для точного проектирования трехмерных каркасных объектов асимметричной и неправильной геометрии, которые будут в центре внимания будущей работы.
Вкратце, алгоритм может обрабатывать произвольные длины ребер и углы вершин для точного проектирования трехмерных каркасных объектов асимметричной и неправильной геометрии, которые будут в центре внимания будущей работы.
Поколение PDB
Программное обеспечение для создания PDB в ATHENA использует узлы уровня оснований нуклеиновых кислот, полученные в результате процедуры маршрутизации, и эти узлы включают информацию о последовательности, маршрутизации и положении каждого основания нуклеиновой кислоты. Первым шагом в создании PDB является маршрутизация информации об узле базового уровня в последовательные нити нуклеиновой кислоты, подходящие для модели всего атома, что выполняется с помощью алгоритма поиска, поскольку каждое основание сопоставляется с вышестоящими, нижестоящими и парными основаниями. в модели.Затем модель всего атома строится по основаниям и по цепям путем преобразования координат эталонной средней базовой структуры нуклеиновой кислоты B-формы в положения узлового уровня. Используемые структуры полностью атомных нуклеиновых кислот взяты из набора параметров 3DNA (29), где координаты основаны на средних структурах ДНК B-формы из Olson et al. (30). Несколько функций преобразования координат ProDy используются во время генерации PDB (31). Области одноцепочечных нуклеиновых кислот не включаются в маршрутизацию на уровне узлов, поэтому непарные координаты интерполируются из ближайших пар оснований выше и ниже по течению с использованием кубической функции Безье, обеспечивая плавный путь от произвольных координат пар оснований.Из-за этой процедуры интерполяции между парами оснований ДНК в вершинах могут возникать стерические конфликты. Для успешного моделирования МД всего атома (32,33) перед стандартными процедурами МД необходима минимизация затронутых участков одноцепочечной ДНК. Предыдущие МД-симуляции всего атома (17, 18, 34) успешно использовали исходные структуры, полученные из DAEDALUS, TALOS и PERDIX соответственно.
Используемые структуры полностью атомных нуклеиновых кислот взяты из набора параметров 3DNA (29), где координаты основаны на средних структурах ДНК B-формы из Olson et al. (30). Несколько функций преобразования координат ProDy используются во время генерации PDB (31). Области одноцепочечных нуклеиновых кислот не включаются в маршрутизацию на уровне узлов, поэтому непарные координаты интерполируются из ближайших пар оснований выше и ниже по течению с использованием кубической функции Безье, обеспечивая плавный путь от произвольных координат пар оснований.Из-за этой процедуры интерполяции между парами оснований ДНК в вершинах могут возникать стерические конфликты. Для успешного моделирования МД всего атома (32,33) перед стандартными процедурами МД необходима минимизация затронутых участков одноцепочечной ДНК. Предыдущие МД-симуляции всего атома (17, 18, 34) успешно использовали исходные структуры, полученные из DAEDALUS, TALOS и PERDIX соответственно.
Стандартный формат файла PDB (22) имеет несколько давних ограничений для больших атомных структур, в том числе ограничения на количество отдельных цепей или нитей нуклеиновой кислоты (62, с учетом количество остатков или оснований нуклеиновых кислот (9999) и пространственные размеры {-999.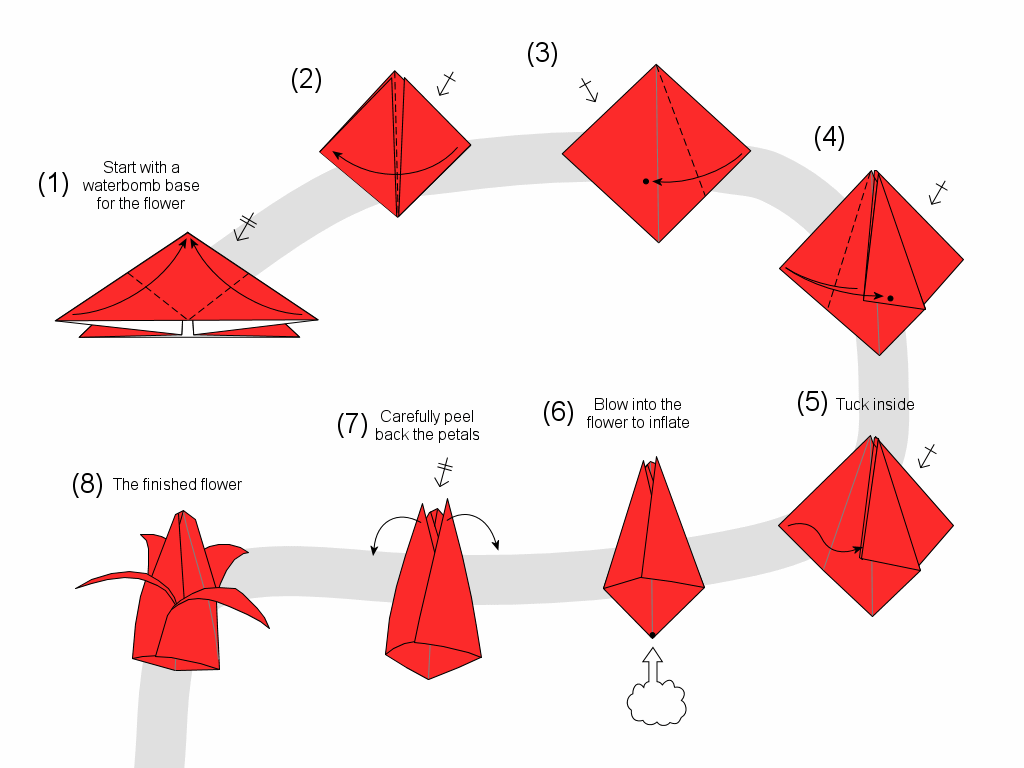 999, 9999,999} в Ангстремах. Это программное обеспечение для создания PDB использует обходные пути для некоторых из этих ограничений. Схема нумерации атомов выше индекса 99 999 использует гибридную схему кодирования с основанием 36, где первый символ является алфавитным с учетом регистра, а следующие четыре символа являются буквенно-цифровыми с основанием 36, что теоретически позволяет использовать более 87 миллионов атомов. Алфавитный первый символ позволяет любому синтаксическому анализатору распознать переход от кодировки base-10 к гибридной кодировке base-36. Схема нумерации остатков над индексом 9999 аналогичным образом допускает >2.Всего 4 миллиона остатков с использованием той же гибридной кодировки base-36. Для более крупных атомных структур, в частности с пространственными размерами, превышающими стандартные ограничения PDB, можно использовать формат файла PDBx/mmCIF (35), но это оставлено для будущей работы. Выходные файлы PDB совместимы с силовыми полями CHARMM для ДНК (36,37), а преобразование в силовые поля AMBER (38) возможно в AmberTools (39).
999, 9999,999} в Ангстремах. Это программное обеспечение для создания PDB использует обходные пути для некоторых из этих ограничений. Схема нумерации атомов выше индекса 99 999 использует гибридную схему кодирования с основанием 36, где первый символ является алфавитным с учетом регистра, а следующие четыре символа являются буквенно-цифровыми с основанием 36, что теоретически позволяет использовать более 87 миллионов атомов. Алфавитный первый символ позволяет любому синтаксическому анализатору распознать переход от кодировки base-10 к гибридной кодировке base-36. Схема нумерации остатков над индексом 9999 аналогичным образом допускает >2.Всего 4 миллиона остатков с использованием той же гибридной кодировки base-36. Для более крупных атомных структур, в частности с пространственными размерами, превышающими стандартные ограничения PDB, можно использовать формат файла PDBx/mmCIF (35), но это оставлено для будущей работы. Выходные файлы PDB совместимы с силовыми полями CHARMM для ДНК (36,37), а преобразование в силовые поля AMBER (38) возможно в AmberTools (39).
Материалы
штапельных нитей ДНК-оригами были приобретены в формате 96-луночных планшетов у Integrated DNA Technologies, Inc.в масштабе синтеза 25 нмоль. Штапельные нити очищали стандартным обессоливанием и калибровали до 200 мкМ на основе полного выхода. Штапельные нити смешивали в равных объемах из соответствующих лунок и использовали непосредственно для фолдинга ДНК-оригами без дополнительной очистки. Цепь ДНК, модифицированная 5′-тиоловым модификатором (C6 S-S), была приобретена у Integrated DNA Technologies, Inc. в масштабе синтеза 100 нмоль со стандартным обессоливанием. Вода, не содержащая нуклеаз, была приобретена у Integrated DNA Technologies, Inc. Каркас ДНК длиной 7249 нуклеотидов (M13mp18) был приобретен у Guild BioSciences в концентрации 100 нМ.Каркас ДНК длиной 2520 нуклеотидов (phPB84) был получен в соответствии с протоколом на основе фага (6). 10-кратный буфер TAE был приобретен у Alfa Aesar. Тетрагидрат ацетата магния (для молекулярной биологии) был приобретен у MilliporeSigma. 1× ТАЕ-буфер с 12,5 мМ Mg(OAc) 2 готовили с 10× ТАЕ-буфером и тетрагидратом ацетата магния. Набор для конъюгации 5 нм OligoREADY AuNP был приобретен у Cytodiagnostics Inc. Pierce DTT (дитиотреитол) был приобретен у ThermoFisher Scientific, а колонки illustra NAP-5 были приобретены у GE Healthcare Life Sciences.
1× ТАЕ-буфер с 12,5 мМ Mg(OAc) 2 готовили с 10× ТАЕ-буфером и тетрагидратом ацетата магния. Набор для конъюгации 5 нм OligoREADY AuNP был приобретен у Cytodiagnostics Inc. Pierce DTT (дитиотреитол) был приобретен у ThermoFisher Scientific, а колонки illustra NAP-5 были приобретены у GE Healthcare Life Sciences.
Оригами для самостоятельной сборки
Все двухмерные пятиугольные ДНК-оригами были сложены по одному и тому же протоколу. 5 нМ каркаса ДНК (M13mp18) смешивали с 20 молярными экв. соответствующие нити скоб в 1× ТАЕ-буфере с 12,5 мМ Mg(OAc) 2 , конечный объем раствора для самосборки составлял 100 мкл. Смесь отжигали в ПЦР-термоциклере: 95°С в течение 2 мин, от 70°С до 45°С со скоростью 0,5°С за 20 мин и от 45°С до 20°С со скоростью 0,5°С за 20 мин. 10 минут.Отожженный раствор был проверен с помощью 1,5% агарозного геля в 1x буфере TAE с 12,5 мМ Mg(OAc) 2 и 1x SybrSafe. Гели запускали при 60 В, а затем визуализировали в синем свете. Отожженный раствор разбавляли до 500 мкл 1× ТАЕ-буфером с 12,5 мМ Mg(OAc) 2 и удаляли лишние основные нити с помощью колонок для концентрации со спин-фильтром MWCO = 100 кДа. Очищенный раствор ДНК оригами доводили до желаемых концентраций (5 нМ) для визуализации с помощью АСМ и ПЭМ.
Отожженный раствор разбавляли до 500 мкл 1× ТАЕ-буфером с 12,5 мМ Mg(OAc) 2 и удаляли лишние основные нити с помощью колонок для концентрации со спин-фильтром MWCO = 100 кДа. Очищенный раствор ДНК оригами доводили до желаемых концентраций (5 нМ) для визуализации с помощью АСМ и ПЭМ.
Объект оригами на основе DX был свернут с использованием каркаса ДНК 40 нМ (phPB84) и 20 молярных эквивалентов необходимых основных цепей в буфере 1× TAE с 12 мМ MgCl 2 .Сворачивающуюся смесь отжигали в термоциклере: 95°С 5 мин, 80°С до 76°С со скоростью 0,8°С/мин, 75°С до 30°С со скоростью 0,42°С/мин. мин и, наконец, 29°С до 25°С со скоростью 0,625°С в мин. Отожженный раствор анализировали в 2,5% агарозном геле в 1x буфере TBE с 12 мМ MgCl 2 и 1x SybrSafe, запускали при 65 В в холодной комнате и визуализировали в синем свете. После удаления скоб и замены буфера на 1× ТАЭ с 8 мМ MgCl 2 с использованием Amicon Ultra 0.Колонки со спин-фильтром объемом 5 мл с MWCO = 100 кДа, очищенный свернутый раствор подвергали скринингу с помощью АСМ и крио-ЭМ визуализации.
Получение конъюгата ДНК-AuNP модифицированной ДНК оригами
5′-тиол-модифицированную цепь ДНК (50 мкМ) восстанавливали с помощью DTT (0,1 М) в 0,15 М натрий-фосфатном буфере (рН 8,5) в течение 2 ч при комнатной температуре. Затем реакционный раствор очищали на колонке Nap-5 для удаления небольших молекул из 5′-цепи тиол-ДНК. Концентрация очищенной 5′-цепи тиол-ДНК была доведена до 25 мкМ в воде, не содержащей нуклеаз, исходя из OD 260 нм .Один флакон с лиофилизированным 5-нм AuNP OligoREADY™ ресуспендировали в 740 мкл не содержащей нуклеазы H 2 O. К AuNP добавляли 160 мкл очищенной 5′-тиоловой цепи ДНК (25 мкМ) и 100 мкл 1 М NaCl приостановка. Смесь инкубировали при комнатной температуре в течение 2 часов. Избыток цепи ДНК затем удаляли из концентрационных колонок с центрифужным фильтром MWCO = 100 кДа, а тем временем конъюгат ДНК-AuNP концентрировали. Концентрацию конъюгата ДНК-AuNP определяли по OD 520 нм .
Конъюгат ДНК-AuNP добавляли к раствору очищенной ДНК оригами (20 нМ) в соотношении 5:1 (AuNP : сайты модификации на оригами), и смеси инкубировали в 1× ТАЕ-буфере с 12,5 мМ Mg( OAc) 2 при комнатной температуре в течение ночи.
АСМ и ПЭМ изображения
АСМ-изображение выполняли в «режиме ScanAsyst в жидкости» (Dimension FastScan, Bruker Corporation) с наконечниками ScanAsyst-Fluid+ или SNL-10 (Bruker Inc.). Два микролитра образца (5 нМ) наносили на свежесколотую слюду (Ted Pella Inc.) и от 0,5 до 1,0 мкл NiCl 2 в концентрации 100 мМ для фиксации наноструктур оригами на поверхности слюды. После ожидания примерно 30 с для адсорбции образца на слюде к образцам добавляли 80 мкл 1× ТАЕ/Mg 2+ буфера, а дополнительные 40 мкл того же буфера наносили на наконечник АСМ. Для визуализации с помощью ПЭМ 5 мкл раствора ДНК-оригами (5 нМ) наносили на свежую углеродную пленку, полученную тлеющим разрядом, с медными сетками (CF200H-CU; Electron Microscopy Sciences Inc.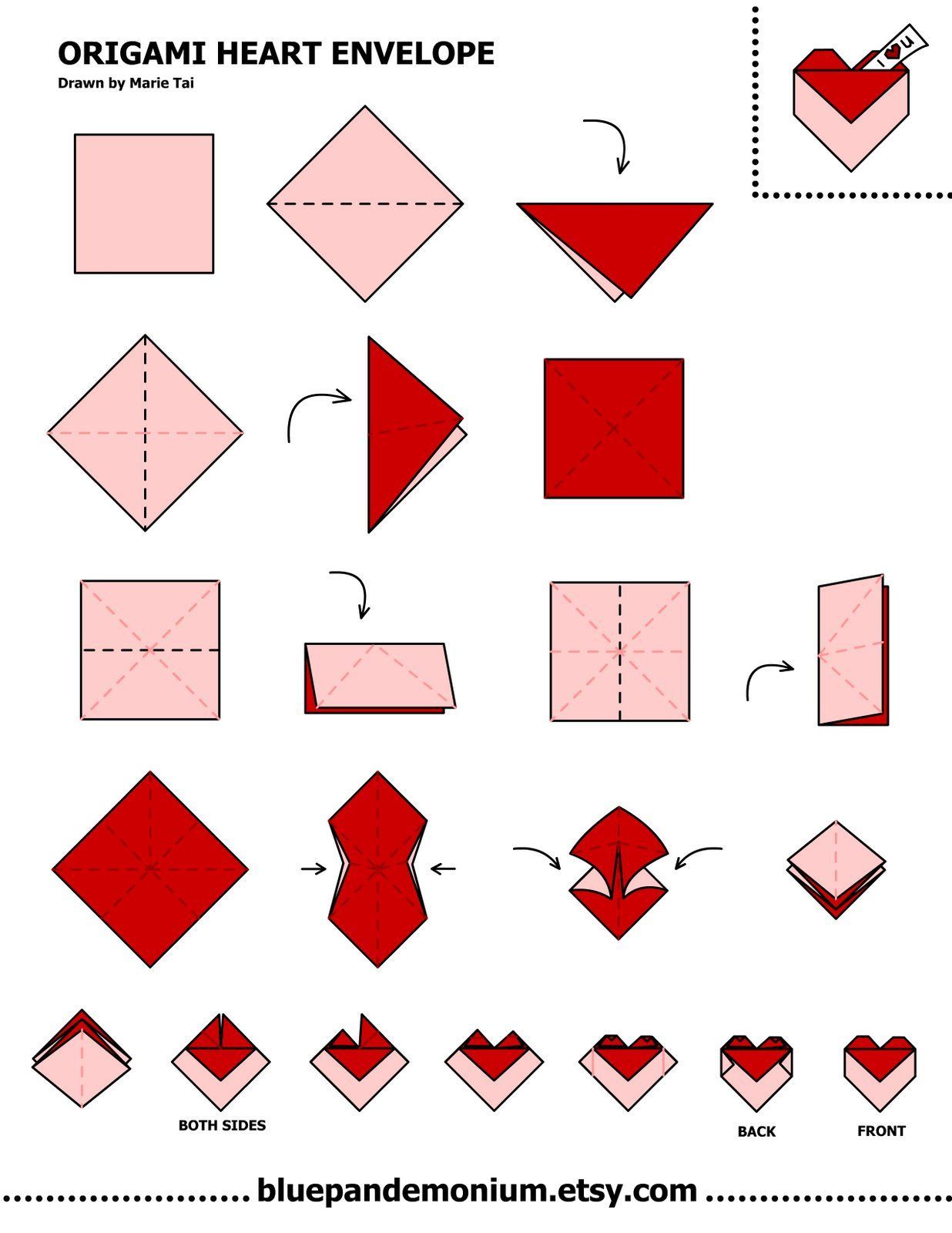 , Hatfeld, PA), после чего образцу давали впитаться в поверхность в течение 30 с. После удаления раствора образца с сетки фильтровальной бумагой Whatman 42 сетку помещали на 5 мкл свежеприготовленного 2%-ного формиата уранила с 25 мМ NaOH на 10 с. Оставшийся раствор красителя на сетке удаляли фильтровальной бумагой Whatman 42 и сушили в домашнем вакууме перед визуализацией. Изображение образца было получено на Tecnai FEI с помощью камеры Gatan.
, Hatfeld, PA), после чего образцу давали впитаться в поверхность в течение 30 с. После удаления раствора образца с сетки фильтровальной бумагой Whatman 42 сетку помещали на 5 мкл свежеприготовленного 2%-ного формиата уранила с 25 мМ NaOH на 10 с. Оставшийся раствор красителя на сетке удаляли фильтровальной бумагой Whatman 42 и сушили в домашнем вакууме перед визуализацией. Изображение образца было получено на Tecnai FEI с помощью камеры Gatan.
Сбор данных крио-ЭМ и обработка изображений
Triton X-100 добавляли к концентрированным очищенным образцам оригами (∼1.3 мкМ) для конечной концентрации 0,025%. Полученный раствор объемом 3 мкл наносили на медные сетки Quantifoil R2/1 с тлеющим разрядом 300 меш и замораживали в жидком этане с помощью Vitrobot (ThermoFisher Scientific) с 6-секундным блоттингом. Затем сетки визуализировали с помощью телескопа Talos Arctica G2 (ThermoFisher Scientific) с детектором Falcon 3EC, работающего при 200 кВ и увеличении 73 000x (номинальный размер пикселя 2,008 Å), с использованием программного обеспечения EPU (ThermoFisher Scientific). Обработка изображения одной частицы и трехмерная реконструкция для асимметричного октаэдра на основе DX с длиной ребра 63 были выполнены с использованием EMAN2 (40).Все частицы были собраны вручную с помощью e2boxer.py в EMAN2. Первоначальные модели, сгенерированные программным обеспечением ATHENA, были отфильтрованы фильтром нижних частот до 60 Å, чтобы избежать систематической ошибки модели. Следующие шаги были выполнены, как описано ранее (17). Всего для окончательного уточнения асимметричного октаэдра на основе DX с длиной ребра 63 п.н. было использовано 3148 частиц.
Обработка изображения одной частицы и трехмерная реконструкция для асимметричного октаэдра на основе DX с длиной ребра 63 были выполнены с использованием EMAN2 (40).Все частицы были собраны вручную с помощью e2boxer.py в EMAN2. Первоначальные модели, сгенерированные программным обеспечением ATHENA, были отфильтрованы фильтром нижних частот до 60 Å, чтобы избежать систематической ошибки модели. Следующие шаги были выполнены, как описано ранее (17). Всего для окончательного уточнения асимметричного октаэдра на основе DX с длиной ребра 63 п.н. было использовано 3148 частиц.
Крупномасштабное компьютерное моделирование с использованием oxDNA
Чтобы продемонстрировать применимость сгенерированных ATHENA ДНК-наноструктур для крупнозернистого моделирования oxDNA, мы выполнили молекулярно-динамическое моделирование (МД) с использованием модели oxDNA2 и версии 2 oxDNA.4 программное обеспечение для моделирования (41–44). По сравнению с полноатомным моделированием МД, oxDNA обеспечивает грубое приближение для изучения термодинамических и механических свойств ДНК в более длительных временных масштабах (41,43). Мы исследовали пентагональные объекты на основе 6HB с различной длиной ребра (от 42 до 210 п.н.) в том виде, в каком они были получены экспериментально. Выходные мультимодельные PDB-файлы ATHENA сначала были преобразованы в файловый формат oxDNA с использованием tacoxDNA (45). Все наноструктуры ДНК были смоделированы при концентрации соли 1 М [Na + ], как было предложено для представления экспериментальных условий (43).После начальной минимизации энергии (2000 шагов) ДНК-объекты уравновешивали в течение 30,3 мкс (10 7 шагов) при 300 К с использованием термостата Ланжевена (коэффициент диффузии 2,5). Затем были смоделированы уравновешенные структуры в течение 0,303 мс (10 8 шагов, временной шаг: 0,1515 пс) при 300 К с использованием Андерсоноподобного термостата Джона (коэффициент диффузии 2,5).
По сравнению с полноатомным моделированием МД, oxDNA обеспечивает грубое приближение для изучения термодинамических и механических свойств ДНК в более длительных временных масштабах (41,43). Мы исследовали пентагональные объекты на основе 6HB с различной длиной ребра (от 42 до 210 п.н.) в том виде, в каком они были получены экспериментально. Выходные мультимодельные PDB-файлы ATHENA сначала были преобразованы в файловый формат oxDNA с использованием tacoxDNA (45). Все наноструктуры ДНК были смоделированы при концентрации соли 1 М [Na + ], как было предложено для представления экспериментальных условий (43).После начальной минимизации энергии (2000 шагов) ДНК-объекты уравновешивали в течение 30,3 мкс (10 7 шагов) при 300 К с использованием термостата Ланжевена (коэффициент диффузии 2,5). Затем были смоделированы уравновешенные структуры в течение 0,303 мс (10 8 шагов, временной шаг: 0,1515 пс) при 300 К с использованием Андерсоноподобного термостата Джона (коэффициент диффузии 2,5). Начальные скорости были сгенерированы из максвелловского распределения. Моделирование визуализировали с помощью oxView и анализировали с помощью инструментов анализа oxDNA (44, 46).Среднеквадратичные колебания (RMSF) рассчитывали относительно средней структуры.
Начальные скорости были сгенерированы из максвелловского распределения. Моделирование визуализировали с помощью oxView и анализировали с помощью инструментов анализа oxDNA (44, 46).Среднеквадратичные колебания (RMSF) рассчитывали относительно средней структуры.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В ATHENA 2D- и 3D-целевые геометрии задаются с помощью полигональной сетки поверхности, а в 3D каждое ребро каждой полигональной поверхности представляет собой одно из ребер соседней поверхности. Они предоставляются вручную или через формат файла ASCII, который определяет полигональную сетку, например, формат файла полигона (PLY), STereoLithography (STL) или язык моделирования виртуальной реальности (WRL) с использованием любого количества программ САПР.Затем ATHENA обеспечивает полностью автоматизированный дизайн последовательности 2D- или 3D-каркаса объектов ДНК-оригами, основанных на жестких 6HB или более совместимых кромках 2HB (рис. 1 и дополнительное примечание 1). Файл PLY (дополнительное примечание 2) используется в качестве входных данных для ATHENA из-за его простоты и широкого применения в проектах на основе САПР. Преобразование типов файлов STL в PLY может выполняться с использованием инструментов с открытым исходным кодом и онлайн-конвертеров. Поскольку маршрутизация скаффолда и дизайн скобок основаны на файлах PLY, важно, чтобы каждая вершина, указанная в файле, относилась хотя бы к одной грани, поскольку в противном случае нет возможности провести скаффолд ssDNA через весь целевой объект (17).После ввода входных данных ATHENA предлагает возможность визуализировать целевой 2D- или 3D-каркасный объект, используя затенение поверхности и/или края каркаса, в цветах по умолчанию, которые можно изменить с помощью пользовательских параметров (рис. 1A; i). Масштабирование, вращение и перевод могут быть выбраны как стандартные параметры мыши, а также перспектива по сравнению с ортогональными видами (рис. 1A; iv). ATHENA также предоставляет 37 предопределенных 2D и 55 3D целевых геометрий (рис.
1 и дополнительное примечание 1). Файл PLY (дополнительное примечание 2) используется в качестве входных данных для ATHENA из-за его простоты и широкого применения в проектах на основе САПР. Преобразование типов файлов STL в PLY может выполняться с использованием инструментов с открытым исходным кодом и онлайн-конвертеров. Поскольку маршрутизация скаффолда и дизайн скобок основаны на файлах PLY, важно, чтобы каждая вершина, указанная в файле, относилась хотя бы к одной грани, поскольку в противном случае нет возможности провести скаффолд ssDNA через весь целевой объект (17).После ввода входных данных ATHENA предлагает возможность визуализировать целевой 2D- или 3D-каркасный объект, используя затенение поверхности и/или края каркаса, в цветах по умолчанию, которые можно изменить с помощью пользовательских параметров (рис. 1A; i). Масштабирование, вращение и перевод могут быть выбраны как стандартные параметры мыши, а также перспектива по сравнению с ортогональными видами (рис. 1A; iv). ATHENA также предоставляет 37 предопределенных 2D и 55 3D целевых геометрий (рис. 1A; ii и дополнительные примечания 3 и 4).
1A; ii и дополнительные примечания 3 и 4).
Рис. 1.
Интерфейс и выходы дизайна ATHENA. ( A ) Снимок экрана графического пользовательского интерфейса с двумя окнами для визуализации целевой геометрии (окно ввода) и выходных данных (окно вывода), таких как цилиндрическая, маршрутная и псевдоатомарная модели. Дополнительные четыре панели предназначены для управления опциями; (i) цветовая схема рендеринга, (ii) целевая геометрия, последовательность каркаса, длина края, тип края, (iii) выходные данные и (iv) управление камерой. ( B ) На основе целевой геометрии DAEDALUS от ATHENA направляет однонитевой каркас по всей геометрии и генерирует несколько выходных данных; цилиндрическая модель, модель маршрутизации, псевдоатомная модель, текстовый файл для основных последовательностей, JSON для caDNAno и PDB для моделирования молекулярной динамики.
Рис. 1.
Интерфейс и выходные данные ATHENA. ( A ) Снимок экрана графического пользовательского интерфейса с двумя окнами для визуализации целевой геометрии (окно ввода) и выходных данных (окно вывода), таких как цилиндрическая, маршрутная и псевдоатомарная модели.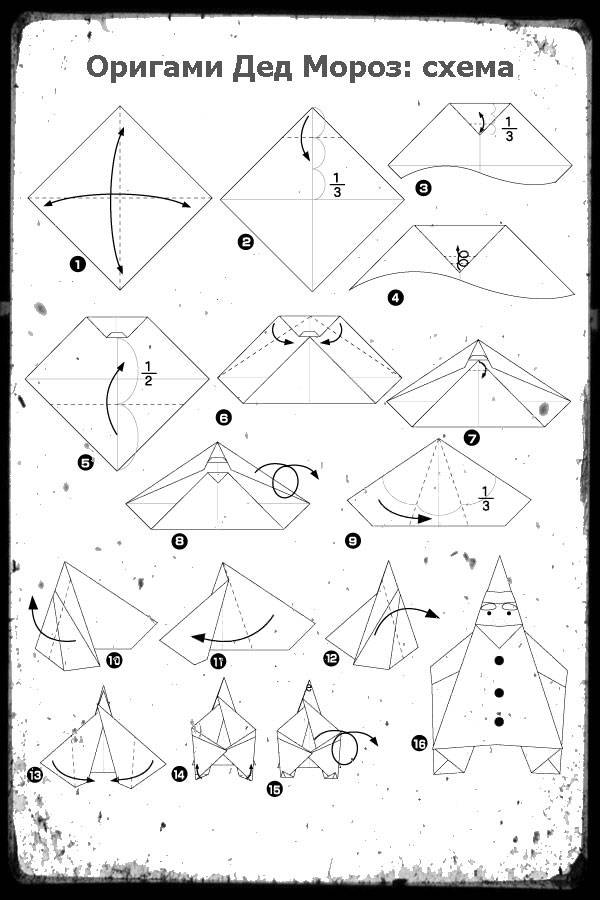 Дополнительные четыре панели предназначены для управления опциями; (i) цветовая схема рендеринга, (ii) целевая геометрия, последовательность каркаса, длина края, тип края, (iii) выходные данные и (iv) управление камерой. ( B ) На основе целевой геометрии DAEDALUS от ATHENA направляет однонитевой каркас по всей геометрии и генерирует несколько выходных данных; цилиндрическая модель, модель маршрутизации, псевдоатомная модель, текстовый файл для основных последовательностей, JSON для caDNAno и PDB для моделирования молекулярной динамики.
Дополнительные четыре панели предназначены для управления опциями; (i) цветовая схема рендеринга, (ii) целевая геометрия, последовательность каркаса, длина края, тип края, (iii) выходные данные и (iv) управление камерой. ( B ) На основе целевой геометрии DAEDALUS от ATHENA направляет однонитевой каркас по всей геометрии и генерирует несколько выходных данных; цилиндрическая модель, модель маршрутизации, псевдоатомная модель, текстовый файл для основных последовательностей, JSON для caDNAno и PDB для моделирования молекулярной динамики.
ATHENA использует M13mp18 в качестве последовательности скаффолда по умолчанию для требуемых длин, меньших или равных 7249 нт; последовательность фага лямбда, если она больше 7250 нуклеотидов и меньше или равна 48502 нуклеотидам; и случайная последовательность, если она больше 48 503 нт. Определяемые пользователем последовательности каркасов также можно импортировать с помощью текстового файла (рис. 1А; ii). В ATHENA есть возможность выбрать тип ребра: двойной кроссовер ДНК (DX или 2HB) или пучок из шести спиралей (6HB), который состоит из каждого ребра 2D- или 3D-каркасных объектов (рис. 1A; ii).Затем можно выполнить полностью автоматизированный дизайн каркаса и последовательности скобок с использованием мотивов DX- (3,16) или 6HB-краев (17,18) либо с каркасом по умолчанию, каркасом одноцепочечной ДНК M13, либо с пользовательским каркасом длины и последовательности, определенными пользователь (рисунок 1A; iii и дополнительный рисунок S1). Кроме того, минимальная длина ребра назначается самому короткому ребру, которое затем используется для масштабирования всех других ребер, определяя длину ребра от 42 п.н. (13,9 нм) до 210 п.н. (71,1 нм) в схеме, что может диапазон от 20 до 200 нм для 2D и от 20 до 100 нм для 3D при использовании каркаса одноцепочечной ДНК M13mp18 (7249 нт).
1A; ii).Затем можно выполнить полностью автоматизированный дизайн каркаса и последовательности скобок с использованием мотивов DX- (3,16) или 6HB-краев (17,18) либо с каркасом по умолчанию, каркасом одноцепочечной ДНК M13, либо с пользовательским каркасом длины и последовательности, определенными пользователь (рисунок 1A; iii и дополнительный рисунок S1). Кроме того, минимальная длина ребра назначается самому короткому ребру, которое затем используется для масштабирования всех других ребер, определяя длину ребра от 42 п.н. (13,9 нм) до 210 п.н. (71,1 нм) в схеме, что может диапазон от 20 до 200 нм для 2D и от 20 до 100 нм для 3D при использовании каркаса одноцепочечной ДНК M13mp18 (7249 нт).
После завершения процедуры проектирования последовательности в ATHENA отображается цилиндрическое представление, перекрывающееся с целевой геометрией (рис. 1B). В цилиндрической модели каждый край каркасной структуры визуализируется с использованием цилиндра (диаметром 2 нм), который представляет собой двойную спираль ДНК.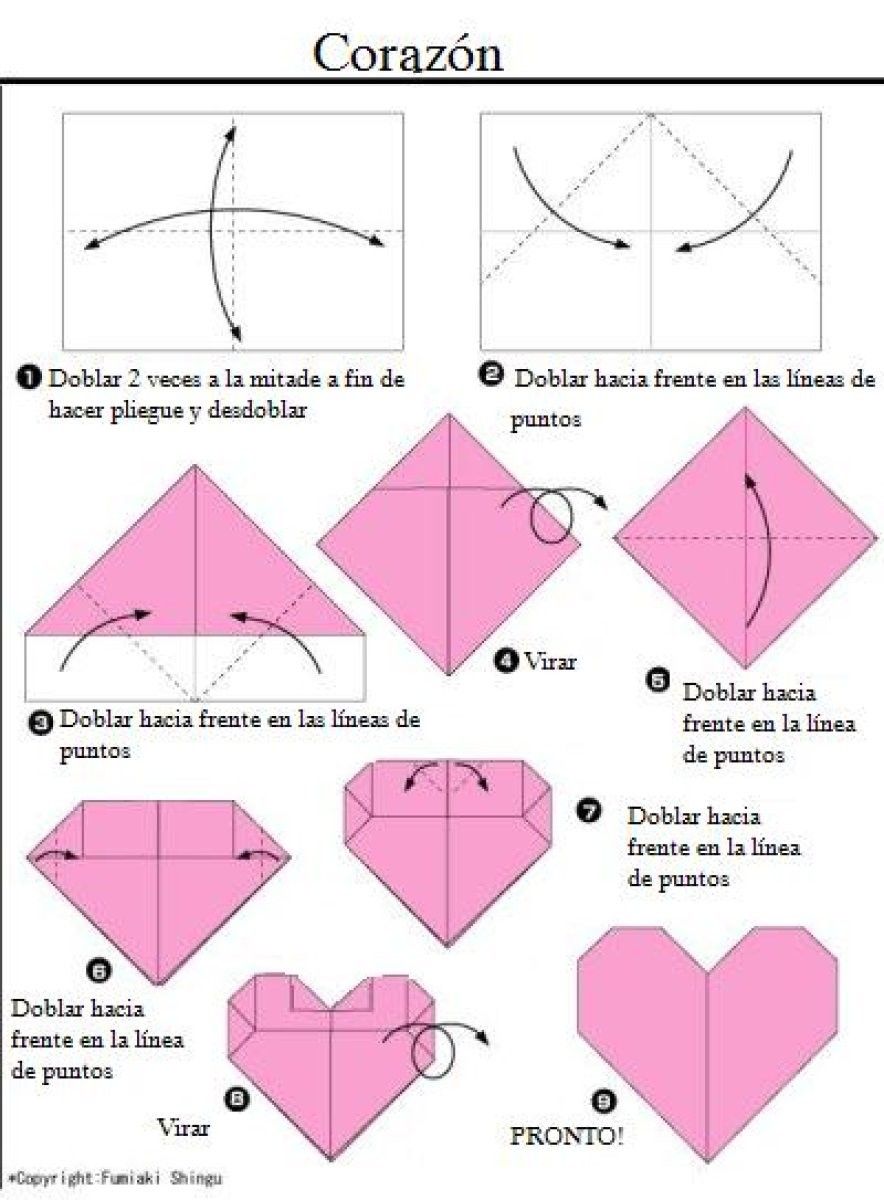 Маршрутизацию нити и спиральность ДНК можно отобразить с помощью параметров маршрутизации и псевдоатомной модели (рис. 1А; iv). Для модели маршрутизации каждая цепь, включая каркас и скобы, аппроксимируется вектором, представляющим направление цепи ДНК (рис. 1В).Более подробные результаты с двойной спиралью ДНК можно отобразить в псевдоатомной модели, построенной сферами и линиями, представляющими нуклеотиды и основу ДНК соответственно. Для облегчения идентификации каркаса и отдельных скоб для маршрутной и псевдоатомарной моделей построены двухцветные схемы с несколькими цветами. Результирующие выходные данные последовательности также можно экспортировать (рис. 1A; iii) в несколько файлов; электронную таблицу значений, разделенных запятыми (CSV), содержащую основные последовательности, модель всего атома PDB и нотацию объектов JavaScript (JSON) для caDNAno (рис. 1B).Веб-сервер tacoxDNA (45) можно использовать для преобразования файла PDB в соответствующие входные данные для выполнения крупномасштабного моделирования с помощью oxDNA (25, 26).
Маршрутизацию нити и спиральность ДНК можно отобразить с помощью параметров маршрутизации и псевдоатомной модели (рис. 1А; iv). Для модели маршрутизации каждая цепь, включая каркас и скобы, аппроксимируется вектором, представляющим направление цепи ДНК (рис. 1В).Более подробные результаты с двойной спиралью ДНК можно отобразить в псевдоатомной модели, построенной сферами и линиями, представляющими нуклеотиды и основу ДНК соответственно. Для облегчения идентификации каркаса и отдельных скоб для маршрутной и псевдоатомарной моделей построены двухцветные схемы с несколькими цветами. Результирующие выходные данные последовательности также можно экспортировать (рис. 1A; iii) в несколько файлов; электронную таблицу значений, разделенных запятыми (CSV), содержащую основные последовательности, модель всего атома PDB и нотацию объектов JavaScript (JSON) для caDNAno (рис. 1B).Веб-сервер tacoxDNA (45) можно использовать для преобразования файла PDB в соответствующие входные данные для выполнения крупномасштабного моделирования с помощью oxDNA (25, 26). Файл JSON можно импортировать в caDNAno (8) для ручного редактирования базы и олигонуклеотидов для функционализации, например, для редактирования последовательностей, удлинения цепей, удаления и добавления нуклеотидов, а также для изменения положения кроссоверов и разрывов. ATHENA предоставляет информацию о краях целевой структуры, связанных с поперечными сечениями в представлении caDNAno (дополнительный рисунок S2).
Файл JSON можно импортировать в caDNAno (8) для ручного редактирования базы и олигонуклеотидов для функционализации, например, для редактирования последовательностей, удлинения цепей, удаления и добавления нуклеотидов, а также для изменения положения кроссоверов и разрывов. ATHENA предоставляет информацию о краях целевой структуры, связанных с поперечными сечениями в представлении caDNAno (дополнительный рисунок S2).
Основываясь только на целевой геометрии, последовательности каркаса и типе кромки (DX или 6HB), встроенные алгоритмы проектирования в ATHENA выполняют автоматическую маршрутизацию каркаса и проектирование последовательности скоб, в дополнение к созданию необходимых скобочных прядей, необходимых для экспериментального складывания конструкции. (Фигура 2). PERDIX выполняет полностью автоматизированную маршрутизацию каркасов и проектирование последовательностей скоб для любой 2D-геометрии произвольной формы с использованием исключительно кромок на основе DX, в то время как METIS проектирует любую 2D-геометрию с использованием механически более жестких сот или краев 6HB. DAEDALUS полностью автоматически решает проблему маршрутизации каркаса и проектирования скоб для любой трехмерной многогранной поверхности, используя исключительно ребра на основе DX, тогда как TALOS визуализирует любую трехмерную многогранную поверхность с использованием механически более жестких сотовых ребер, что также требует большей длины каркаса для той же геометрии и размера частиц. TALOS дополнительно предлагает возможность использовать каждый возможный кроссовер между соседними дуплексами ребер 6HB (17), что должно обеспечивать повышенную механическую и ферментативную целостность по сравнению с минимальным количеством одиночных кроссоверов, используемых между любыми двумя ребрами в предыдущих конструкциях сотовых октаэдрических последовательностей (47).
DAEDALUS полностью автоматически решает проблему маршрутизации каркаса и проектирования скоб для любой трехмерной многогранной поверхности, используя исключительно ребра на основе DX, тогда как TALOS визуализирует любую трехмерную многогранную поверхность с использованием механически более жестких сотовых ребер, что также требует большей длины каркаса для той же геометрии и размера частиц. TALOS дополнительно предлагает возможность использовать каждый возможный кроссовер между соседними дуплексами ребер 6HB (17), что должно обеспечивать повышенную механическую и ферментативную целостность по сравнению с минимальным количеством одиночных кроссоверов, используемых между любыми двумя ребрами в предыдущих конструкциях сотовых октаэдрических последовательностей (47).
Рис. 2.
Полностью автоматизированное проектирование последовательности 2D- и 3D-каркасных структур ДНК с ребрами на основе DX и 6HB. Масштабные линейки, 50 и 150 нм (увеличенные и уменьшенные изображения АСМ соответственно) для структуры PERDIX (16), 50 и 200 нм (увеличенные и уменьшенные изображения АСМ соответственно) для структуры METIS (18) и 20 нм (крио-ЭМ-изображение) и 5 нм (крио-ЭМ-реконструкция) для DAEDALUS, расширенного в этой работе, и структур TALOS (3,17).
Рис. 2.
Полностью автоматизированное проектирование последовательности 2D- и 3D-каркасных структур ДНК с ребрами на основе DX и 6HB.Масштабные линейки, 50 и 150 нм (увеличенные и уменьшенные изображения АСМ соответственно) для структуры PERDIX (16), 50 и 200 нм (увеличенные и уменьшенные изображения АСМ соответственно) для структуры METIS (18) и 20 нм (крио-ЭМ-изображение) и 5 нм (крио-ЭМ-реконструкция) для DAEDALUS, расширенного в этой работе, и структур TALOS (3,17).
Мы протестировали способность ATHENA генерировать высококачественные каркасные ДНК-структуры оригами, что также позволяет пользователям удобно функционализировать такие структуры с помощью других материалов.Во-первых, чтобы оценить способность ATHENA обрабатывать ребра произвольной длины для асимметричных и нерегулярных объектов на основе DX-ребер, мы разработали асимметричный октаэдр с постоянной длиной ребра и переменными углами при вершинах с ребрами 2HB. Мы использовали введенную пользователем последовательность (phPB84 с 2520 нуклеотидами, дополнительные таблицы S1 и S2) в качестве каркаса. АСМ и крио-ЭМ подтвердили успешную сборку этой структуры с высоким выходом (рисунок 3 и дополнительный рисунок S3), предложив первую демонстрацию этого нового алгоритма проектирования последовательности.Крио-ЭМ-реконструкция асимметричного октаэдра на основе DX с длиной ребра 63 п.н. также показала, что эти неправильные объекты складываются, как предполагалось, без значительных искажений запрограммированных ребер или вершин (рис. 3E).
АСМ и крио-ЭМ подтвердили успешную сборку этой структуры с высоким выходом (рисунок 3 и дополнительный рисунок S3), предложив первую демонстрацию этого нового алгоритма проектирования последовательности.Крио-ЭМ-реконструкция асимметричного октаэдра на основе DX с длиной ребра 63 п.н. также показала, что эти неправильные объекты складываются, как предполагалось, без значительных искажений запрограммированных ребер или вершин (рис. 3E).
Рисунок 3.
Проектирование асимметричных и неправильных объектов на основе DX с помощью ATHENA. ( A ) Целевая геометрия и модель маршрутизации асимметричного октаэдра с длиной ребра 63 п.н. ( B ) Электрофорез в агарозном геле для асимметричного октаэдра на основе DX.( C ) АСМ и ( D ) крио-ЭМ изображения асимметричного октаэдра на основе DX с длиной ребра 63 п.н. ( E ) Трехмерная реконструкция асимметричного октаэдра на основе DX с длиной ребра 63 п. н. с использованием крио-ЭМ. Разрешение решаемой структуры составляет 3,3 нм.
н. с использованием крио-ЭМ. Разрешение решаемой структуры составляет 3,3 нм.
Рис. 3.
Проектирование асимметричных и нерегулярных объектов на основе DX с помощью ATHENA. ( A ) Целевая геометрия и модель маршрутизации асимметричного октаэдра с длиной ребра 63 п.н. ( B ) Электрофорез в агарозном геле для асимметричного октаэдра на основе DX.( C ) АСМ и ( D ) крио-ЭМ изображения асимметричного октаэдра на основе DX с длиной ребра 63 п.н. ( E ) Трехмерная реконструкция асимметричного октаэдра на основе DX с длиной ребра 63 п.н. с использованием крио-ЭМ. Разрешение решаемой структуры составляет 3,3 нм.
Затем мы создали последовательности основных цепей пяти пятиугольных объектов на основе 6HB (рис. 4 и дополнительные таблицы S3–S8) с различной длиной края от 42 п.н. (13,94 нм) до 210 п.н. (71,06 нм) с помощью ATHENA. ПЭМ и АСМ подтвердили успешную сборку целевых структур, на что указывают точные углы при вершине и высокий выход правильного формирования этих структур (дополнительные рисунки S4 – S13).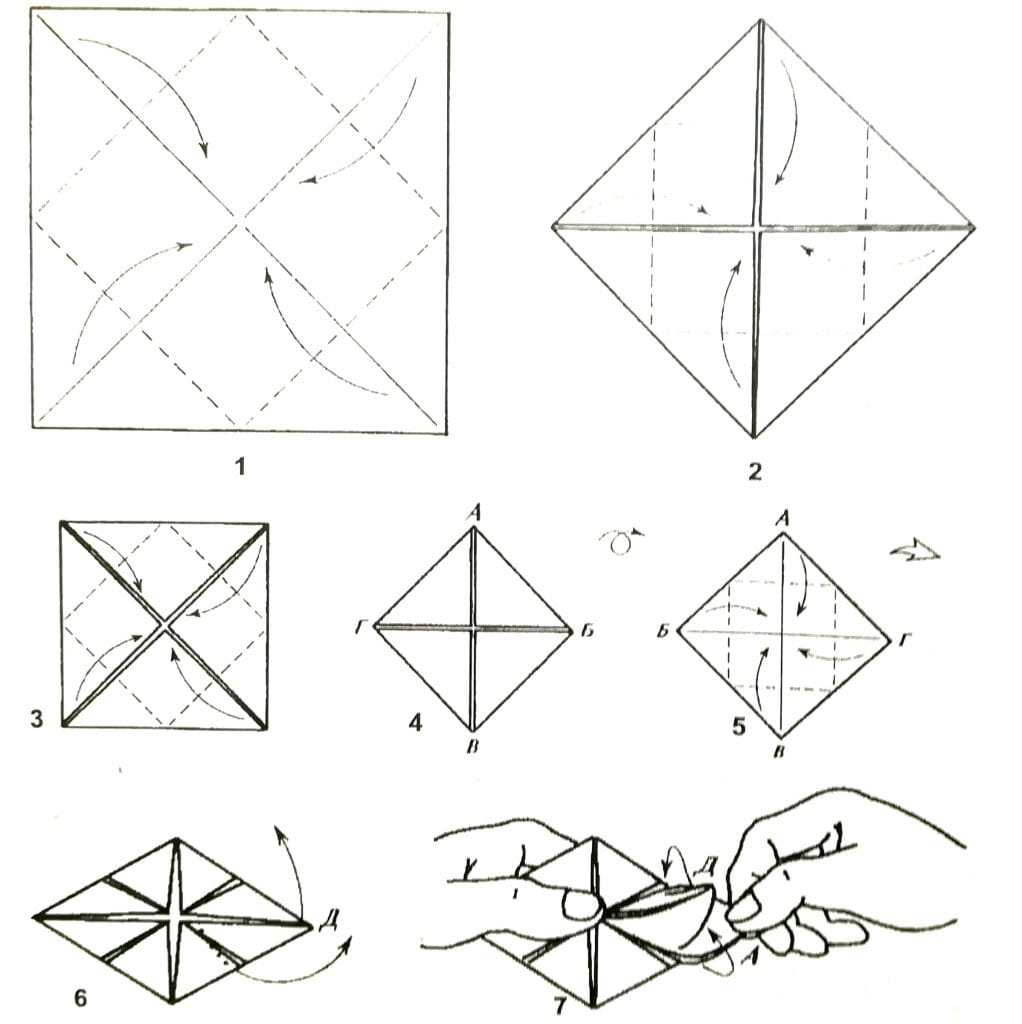 Крупнозернистое моделирование oxDNA подтвердило высокую структурную стабильность и жесткость структур, особенно с меньшей длиной края (рис. 4). Пользователи могут изменять эти структуры на основе трассировки и псевдоатомарной модели, созданной ATHENA, что позволяет пользователю идентифицировать положение конкретной модификации (выемка или положение выступа). Каждая основная нить была помечена одним и тем же цветом как в псевдоатомной модели, так и в файле caDNAno для удобства идентификации соответствующих основных цепей в файле caDNAno для модификаций.Чтобы продемонстрировать адресность этого хорошо контролируемого материала каркаса и подхода к редактированию, мы модифицировали одну из пятиугольных структур оригами (длина ребра 210 п.н.) для прикрепления AuNP (рис. 5). Следуя процедуре, описанной на дополнительном рисунке S14, мы модифицировали основные нити вокруг вершины этой пятиугольной структуры для позиционирования AuNP. Ручки для конъюгатов ДНК-AuNP располагали либо в трех, либо во всех пяти вершинах пентагональной структуры, а ручки от соседних ребер предназначались для фиксации одной AuNP в вершине.
Крупнозернистое моделирование oxDNA подтвердило высокую структурную стабильность и жесткость структур, особенно с меньшей длиной края (рис. 4). Пользователи могут изменять эти структуры на основе трассировки и псевдоатомарной модели, созданной ATHENA, что позволяет пользователю идентифицировать положение конкретной модификации (выемка или положение выступа). Каждая основная нить была помечена одним и тем же цветом как в псевдоатомной модели, так и в файле caDNAno для удобства идентификации соответствующих основных цепей в файле caDNAno для модификаций.Чтобы продемонстрировать адресность этого хорошо контролируемого материала каркаса и подхода к редактированию, мы модифицировали одну из пятиугольных структур оригами (длина ребра 210 п.н.) для прикрепления AuNP (рис. 5). Следуя процедуре, описанной на дополнительном рисунке S14, мы модифицировали основные нити вокруг вершины этой пятиугольной структуры для позиционирования AuNP. Ручки для конъюгатов ДНК-AuNP располагали либо в трех, либо во всех пяти вершинах пентагональной структуры, а ручки от соседних ребер предназначались для фиксации одной AuNP в вершине. (Рисунок 5C и дополнительные рисунки S15-S17). Изображения ПЭМ показали, что AuNP были успешно размещены в заданных позициях в структуре оригами, что альтернативно могло быть использовано для программирования любого количества неорганических или органических молекул как в 2D, так и в 3D (19).
(Рисунок 5C и дополнительные рисунки S15-S17). Изображения ПЭМ показали, что AuNP были успешно размещены в заданных позициях в структуре оригами, что альтернативно могло быть использовано для программирования любого количества неорганических или органических молекул как в 2D, так и в 3D (19).
Рис. 4.
Создание пятиугольных ДНК-оригами из 6HB с ребрами разной длины. ( A ) результаты моделирования oxDNA (центроидные структуры в окраске RMSF) и ( B ) изображения ПЭМ и АСМ для различных длин ребер пятиугольных объектов ДНК длиной 42, 84, 126, 168 и 210 п.н.Масштабные линейки, 25 и 100 нм (увеличение и уменьшение изображения ПЭМ и АСМ соответственно).
Рис. 4.
Создание пятиугольных ДНК-оригами из 6HB с ребрами разной длины. ( A ) результаты моделирования oxDNA (центроидные структуры в окраске RMSF) и ( B ) изображения ПЭМ и АСМ для различных длин ребер пятиугольных объектов ДНК длиной 42, 84, 126, 168 и 210 п.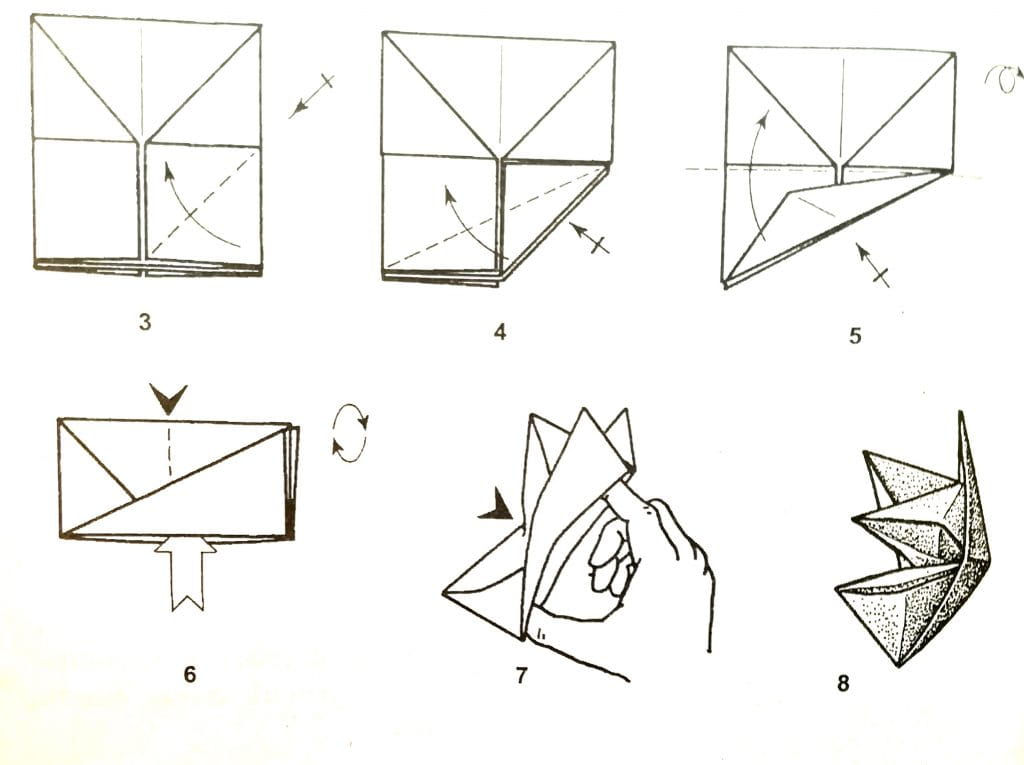 н. Масштабные линейки, 25 и 100 нм (увеличение и уменьшение изображения ПЭМ и АСМ соответственно).
н. Масштабные линейки, 25 и 100 нм (увеличение и уменьшение изображения ПЭМ и АСМ соответственно).
Рисунок 5.
Организация AuNP на пентагональной ДНК-оригами длиной 210 п.н. ( A ) Схемы, показывающие крепление NP в трех или пяти углах. ( B ) Модель маршрутизации и представление caDNAno для пентагональной ДНК-оригами с AuNP. ( C ) ПЭМ-изображения с 2D-оригами пятиугольной ДНК организованных AuNP. Шкала баров, 50 нм.
Рис. 5.
Организация AuNP на пентагональной ДНК-оригами длиной 210 п.н. ( A ) Схемы, показывающие крепление NP в трех или пяти углах.( B ) Модель маршрутизации и представление caDNAno для пентагональной ДНК-оригами с AuNP. ( C ) ПЭМ-изображения с 2D-оригами пятиугольной ДНК организованных AuNP. Шкала баров, 50 нм.
В совокупности предыдущие результаты показывают, что ATHENA предлагает унифицированную программную среду для полностью автоматического нисходящего геометрического проектирования произвольных каркасных конструкций оригами, включая впервые полностью асимметричные каркасные конструкции 2HB, реализованные с помощью нового алгоритма, который обеспечивает непрерывную кромку. длины и произвольные углы вершин, которые нельзя обработать с помощью оригинальной версии DAEDALUS (3).Объекты, сложенные и модифицированные с использованием моделей, созданных ATHENA, демонстрируют надежность и простоту использования этого интерфейса, предлагая универсальный инструмент проектирования для широкого спектра 2D- и 3D-каркасных объектов ДНК-оригами.
длины и произвольные углы вершин, которые нельзя обработать с помощью оригинальной версии DAEDALUS (3).Объекты, сложенные и модифицированные с использованием моделей, созданных ATHENA, демонстрируют надежность и простоту использования этого интерфейса, предлагая универсальный инструмент проектирования для широкого спектра 2D- и 3D-каркасных объектов ДНК-оригами.
ДОПОЛНИТЕЛЬНЫЕ ДАННЫЕ
Дополнительные данные доступны на сайте NAR Online.
ФИНАНСИРОВАНИЕ
Национальный научный фонд [CCF-1956054, CCF-1564025, CBET-1729397]; Управление военно-морских исследований [N00014-17-1-2609, N00014-13-1-0664, N00014-16-1-2506]; Армейская субнаграда ICB [KK1954]; Т.Дж. выражает благодарность Фонду Александра фон Гумбольдта за финансовую поддержку через исследовательскую стипендию Федора-Лайнена. Финансирование платы за открытый доступ: Национальный научный фонд [CCF-1956054, CCF-1564025, CBET-1729397]; Управление военно-морских исследований [N00014-17-1-2609, N00014-13-1-0664, N00014-16-1-2506]; Армейская субнаграда ICB [KK1954].
Заявление о конфликте интересов . Ни один не заявил.
Примечания
Текущий адрес: Уильям П. Брикер, кафедра химической и биологической инженерии, Университет Нью-Мексико, Альбукерке, Нью-Мексико, 87131, США.
ССЫЛКИ
1.Seeman
N.C.
Соединения и решетки нуклеиновых кислот
.Ж. Теор. биол.
1982
;99
:237
–247
.2.Ротемунд
П.В.К.
Сворачивание ДНК для создания наноформ и узоров
.Природа
.2006
;440
:297
–302
.3.Венециано
Р.
,RATANALERT
S.
,S.
,ZHANG
K.
,K.
,ZHANG
F.
,yan
H.
,CHIU
W.
,CARTE
M.
Дизайнерские наноразмерные сборки ДНК, запрограммированные сверху вниз
.
Наука
.2016
;352
:1534
–1534
.4.Veneziano
R.
,Shepherd
T.R.
,RATANALERT
,S.
,S.
,BELLU
L.
,TAO
C.
,
C.
,
CARTE
M.
in vitro Синтез одноцепочечной ДНК ДНК на
.науч. Респ.
2018
;8
:6548
.5.Praetorius
F.
,Kick
B.
,Behler
K.L.
,Хонеманн
М.Н.
,Веустер-Ботц
Д.
,Dietz
H.
Биотехнологическое массовое производство ДНК оригами
.Природа
.2017
;552
:84
–87
.6.Пастух
Т.Р.
,Du
R.R.
,Huang
H.
,Wamhoff
E.-C.
,Bathe
M.
Биопродукция чистой одноцепочечной ДНК размером в тысячу оснований
.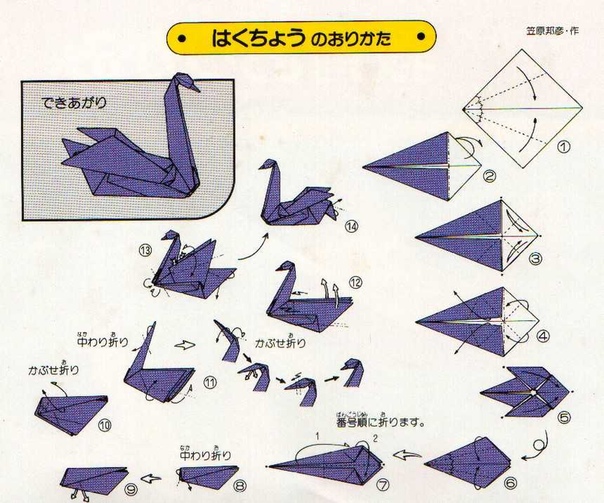
науч. Респ.
2019
;9
:6121
.7.Дуглас
С.М.
,Dietz
H.
,LEDL
T.
,Högberg
,B.
,B.
,GRAF
F.
,SHIH
Вт.м.
Самосборка ДНК в наноразмерные трехмерные формы
.Природа
.2009
;459
:414
–418
.8.Дуглас
С.М.
,Мраморный камень
А.Х.
,Тирапиттаянон
С.
,Васкес
А.
,Церковь
Г.М.
,Ши
В.М.
Быстрое прототипирование трехмерных ДНК-оригами с помощью caDNAno
.Нуклеин. Кислоты. Рез.
2009
;37
:5001
–5006
.9.Дитц
Х.
,Дуглас
С.М.
,Ши
В.М.
Сворачивание ДНК в скрученные и изогнутые наноформы
.Наука
.2009
;325
:725
–730
. 10.
10.Castro
C.E.
,Kilchherr
F.
,Kim
D.-N.
,Шиао
Э.Л.
,Wauer
T.
,Wortmann
P.
,Bathe
M.
,Dietz
H.
Праймер для ДНК-оригами
.Нац. Методы
.2011
;8
:221
–229
.11.Benson
E.
,Mohammed
A.
,Gardell
J.
,Masich
S.
,CCEIZLER
E.
,ORPONEN
стр.
,Högberg
B.
Визуализация ДНК многогранных сеток в наномасштабе
.Природа
.2015
;523
:441
–444
.12.Benson
E.
,Mohammed
A.
,Bosco
A.
,Teixeira
A.I.
,Orponen
P.
,Högberg
B.
Компьютерное производство каркасных наноструктур ДНК из плоских сеток
.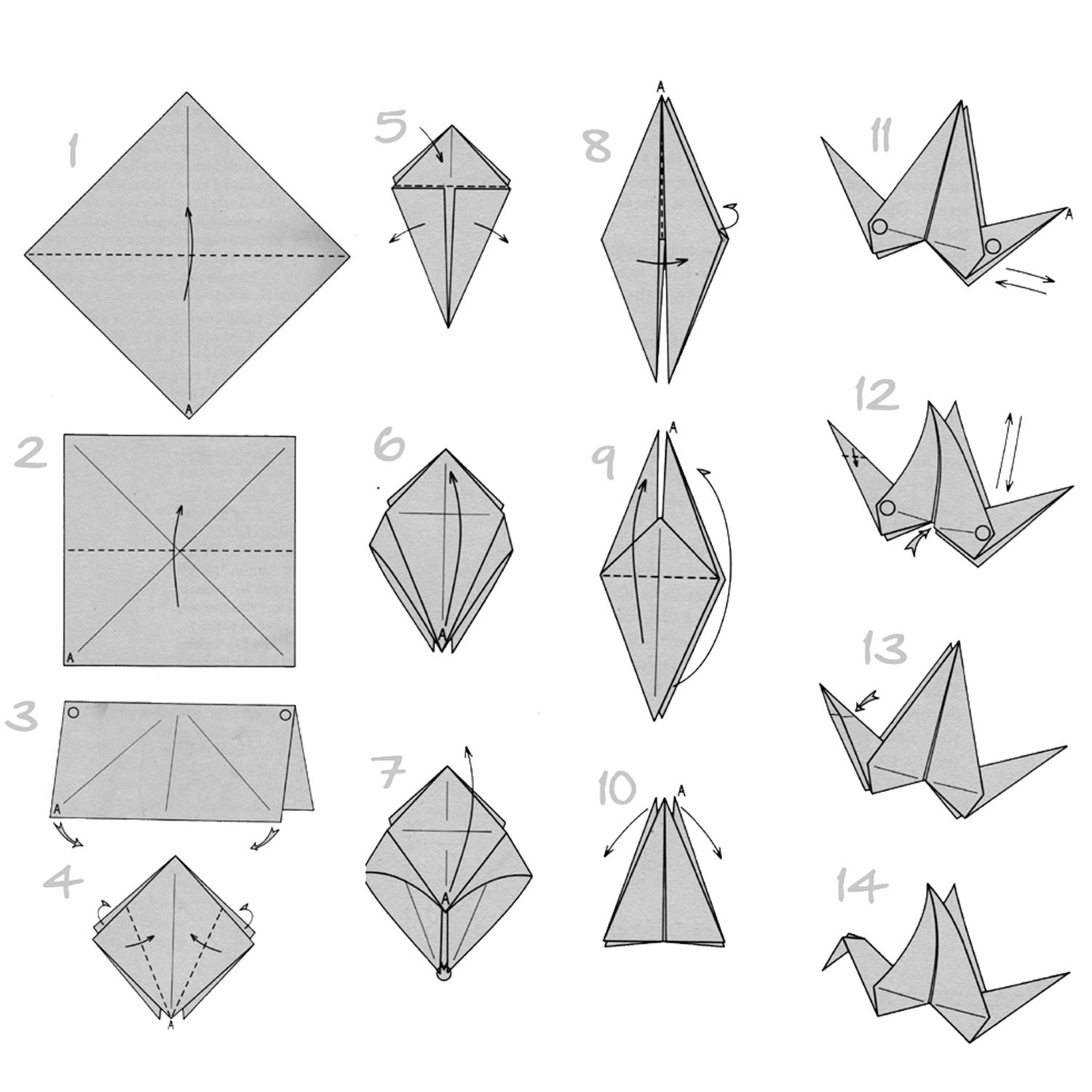
Анжю. хим. Междунар. Эд.
2016
;55
:8869
–8872
.13.Мэттис
М.
,Агарвал
Н.П.
,Шмидт
Т.Л.
Дизайн и синтез триангулированных связок ДНК оригами
.Нано Летт.
2016
;16
:2108
–2113
.14.Ян
Х.
,Парк
С.Х.
,Финкельштейн
Г.
,Райф
Дж.Х.
,ЛаБеан
Т.H.
Самосборка белковых массивов и высокопроводящих нанопроводов на основе ДНК-шаблона
.Наука
.2003
;301
:1882
–1884
.15.Чжан
F.
,jiang
S.
,Â
,
,
S.
,LI
Y.
,MAO
C.
,LIU
Y.
,Yan
H.
Сложные каркасные ДНК-оригами наноструктуры с вершинами многоплечевого соединения
. Нац. нанотехнологии.
2015
;10
:779
–784
.16.июня
г.
,Чжан
F.
,F.
,T.
,,
S.
,qi
x.
,yan
H.
,Bathe
M.
Автономно разработанное двухмерное ДНК-оригами произвольной формы
.науч. Доп.
2019
;5
:eaav0655
.17.Июн
Х.
,Пастух
Т.Р.
,Чжан
К.
,Брикер
В.П.
,Li
S.
,Chiu
W.
,Bathe
M.
Автоматизированное проектирование последовательности трехмерных многогранных каркасных ДНК-оригами 09 909
АКС Нано
.2019
;13
:2083
–2093
.18.Июн
Х.
,Wang
X.
,William
B.
,Bathe
M.
Автоматизированный дизайн последовательности двухмерного каркаса ДНК оригами с 0009 ребрами.
Нац. коммун.
2019
;10
:5419
.19.Вамхофф
Э.-К.
,Banal
J.
,Bricker
W.
,Shepherd
T.
,Parsons
M.
,Veneziano
R.
,R.
,Conee
M.
,М.
,июня
H.
,Wang
x.
,
CARTE
M.
Программирование структурированных ДНК Сборки для зондового биофизического процессы
.год. Преподобный Биофиз.
2019
;48
:395
–419
.20.Венециано
Р.
,Мойер
Т.Дж.
,Камень
М.Б.
,Вамхофф
Э.-К.
,Рид
Б.Дж.
,Мукерджи
С.
,Шеперд
Т.Р.
,Das
J.
,Schief
W.R.
,Irvine
D.J.
и др. .Роль наноразмерной организации антигена в активации В-клеток, исследованная с использованием ДНК-оригами
.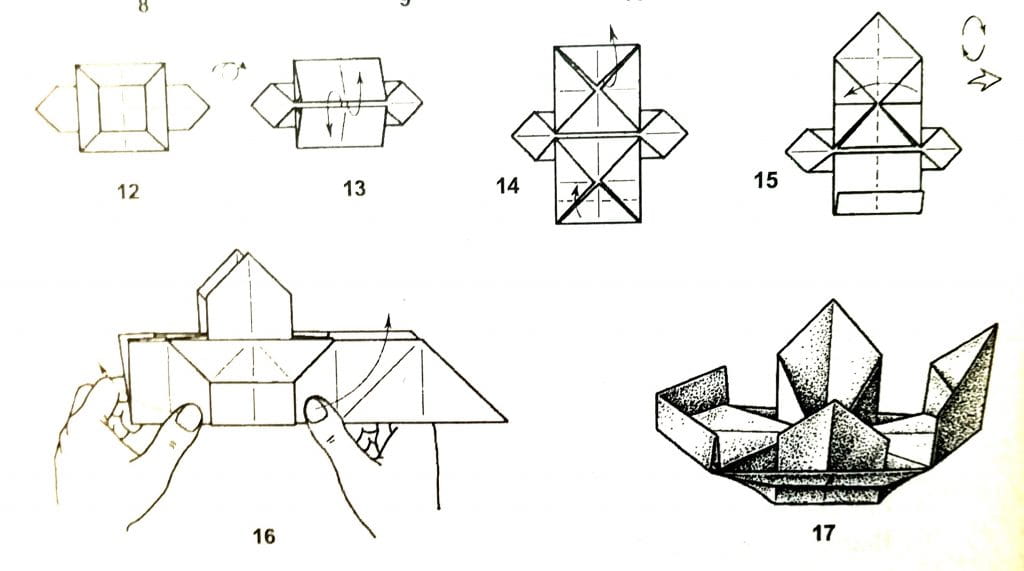
Нац. нанотехнологии.
2020
;15
:716
–723
.21.де Льяно
Э.
,Мяо
Х.
,Ахмади
Ю.
,Уилсон
А.Дж.
,Beeby
M.
,Viola
I.
,Barisic
I.
Adenita: интерактивное 3D моделирование и визуализация наноструктур ДНК 9009 .
Нуклеин. Кислоты. Рез.
2020
;48
:8269
–8275
.22.Берман
Х.M.
,Westbrook
J.
,Feng
Z.
,Gilliland
G.
,Bhat
T.N.
,Вайсиг
Х.
,Шиндялов
И.Н.
,Борн
Ч.П.
Банк данных о белках
.Рез. нуклеиновых кислот.
2000
;28
:235
–242
.23.Хамфри
В.
,Далке
А.
,Schulten
K.
VMD: визуальная молекулярная динамика
.
Дж. Мол. График
1996
;14
:33
–38
.24.Петтерсен
Э.Ф.
,Годдард
Т.Д.
,Хуан
К.С.
,Диван
Г.С.
,Гринблатт
Д.М.
,Мэн
ЕС
,Феррин
Т.Е.
Химера UCSF — система визуализации для поисковых исследований и анализа
.Дж. Вычисл. хим.
2004
;25
:1605
–1612
.25.Оулдридж
Т.Е.
,Луи
А.А.
,Дойе
J.P.K.
Структурные, механические и термодинамические свойства крупнозернистой модели ДНК
.J. Chem. физ.
2011
;134
:085101
.26.Снодин
Б.Е.К.
,Рандиси
Ф.
,Мосайеби
М.
,Шульц
П.
,Шрек
Дж.С. ,Романо
Ф.
,Олдридж
Т.Е.
,Цуканов
Р.
,Нир
Э.
,Луис
А.А.
и др. .Внедрение улучшенных структурных свойств и солевой зависимости в крупнозернистую модель ДНК
.J. Chem. физ.
2015
;142
:234901
.27.Maffeo
C.
,Aksimentiev
A.
MrDNA: модель с несколькими разрешениями для предсказания структуры и динамики систем ДНК
.Рез. нуклеиновых кислот.
2020
;48
:5135
–5146
.28.Ли
Дж.Ю.
,Ли
Дж.Г.
,Юн
Г.
,Ли
К.
,Ким
Ю.-Дж.
,Ким
К.С.
,Ким
Т.Х.
,Ким
Д.-Н.
Быстрый вычислительный анализ сборок ДНК-оригами с разрешением, близким к атомному
.АКС Нано
.2021
;15
:1002
–1015
.29.Лу
X.-J.
3DNA: пакет программного обеспечения для анализа, реконструкции и визуализации трехмерных структур нуклеиновых кислот
.Рез. нуклеиновых кислот.
2003
;31
:5108
–5121
.30.Олсон
В.К.
,Бансал
М.
,Берли
С.К.
,Дикерсон
Р.Е.
,Gerstein
M.
,Harvey
S.C.
,Нейдл
С.
,Шакед
З.
и др. .Стандартная система отсчета для описания геометрии пар оснований нуклеиновых кислот
.Дж. Мол. биол.
2001
;313
:229
–237
.31.Bakan
A.
,Meireles
LM
,Bahar
I.
ProDy: динамика белков, выведенная из теории и экспериментов 9 .
Биоинформатика
.2011
;27
:1575
–1577
.32.Yoo
J.
,Aksimentiev
A.
In situ структура и динамика ДНК-оригами, определенная с помощью моделирования молекулярной динамики
.Проц. Натл. акад. науч. США
2013
;110
:20099
–20104
.33.Maffeo
C.
,Yoo
J.
,Аксиментьев
A.
Реконструкция ДНК оригами структур de novo с помощью моделирования молекулярной динамики 90 09 .
Рез. нуклеиновых кислот.
2016
;44
:3013
–3019
.34.Кастрюля
К.
,Брикер
В.П.
,Ratanalert
S.
,Bathe
M.
Структура и конформационная динамика наночастиц ДНК — оригами
.Рез. нуклеиновых кислот.
2017
;45
:6284
–6298
.35.Вестбрук
Дж.Д.
,Фицджеральд
П.М.Д.
Формат PDB, форматы mmCIF и другие форматы данных
.Структурная биоинформатика
.2005
;John Wiley & Sons, Ltd
159
–179
.36.Foloppe
N.
,A.D.M.
JrПолноатомное эмпирическое силовое поле для нуклеиновых кислот. I. Оптимизация параметров на основе данных о малых молекулах и макромолекулярных мишенях в конденсированной фазе
.Дж. Вычисл. хим.
2000
;21
:86
–104
.37.МакКерелл
Н.Э.
,Банавали
Н.К.
Полноатомное эмпирическое силовое поле для нуклеиновых кислот: II. Приложение к моделированию молекулярной динамики ДНК и РНК в растворе
.Дж. Вычисл. хим.
2000
;21
:105
–120
.38.Галиндо-Мурильо
Р.
,robertson
j.c.
,Zgarbová
,Zgarbová
м.
,ŠPONER
J.
,J.
,,
М.
,Jurečka
стр.
,Chatham
T.E.
Оценка текущего состояния модификаций янтарного силового поля для ДНК
.J. Chem. Теория вычисл.
2016
;12
:4114
–4127
.39.Кейс
D.А.
,Актульга
Х.М.
,Белфон
К.
,Бен-Шалом
И.Ю.
,Брозелл
С.Р.
,Cerutti
D.S.
,Cheatham
T.E.
III,Крузейро
V.W.D.
,Дарден
Т.А.
,Герцог
Р.Э.
и др. .Янтарный 2021
.2021
;Сан-Франциско
Калифорнийский университет
.40.Tang
G.
,PENG
L.
,,
PR
,MANN
DS
,DS
,Jiang
W.
,Rees
I.
,Людтке
SJ
EMAN2: расширяемый пакет обработки изображений для электронной микроскопии
.Дж. Структура. биол.
2007
;157
:38
–46
.41.Шульц
П.
,Романо
Ф.
,Олдридж
Т.Е.
,Rovigatti
L.
,Doye
J.P.K.
,Луи
А.А.
Зависимая от последовательности термодинамика крупнозернистой модели ДНК
.J. Chem. физ.
2012
;137
:135101
.42.Ровигатти
Л.
,Шульц
П.
,Регул
И.Z.
,Romano
F.
Сравнение подходов к распараллеливанию в моделировании молекулярной динамики на графических процессорах
.Дж. Вычисл. хим.
2015
;36
:1
–8
.43.Дойе
J.P.K.
,Fowler
H.
,Prešern
D.
,Bohlin
J.
,Rovigatti
L.
,Romano
F.
,Шульц
П.
,Вонг
К.К.
,Луи
А.А.
,Шрек
Дж.С.
и др. .Крупнозернистая модель oxDNA как инструмент для моделирования ДНК-оригами
.2020
; https://arxiv.org/abs/2004.05052.44.POPPLETON
E.
,RoMero
R
,R.
,Mallya
A.
,Rovigatti
L.
,ŠULC
P.
OxDNA.org: общедоступный веб-сервер для грубого моделирования наноструктур ДНК и РНК
.Рез. нуклеиновых кислот.
2021
;49
:W491
–W498
.45.SUMA
A.
,A.
,E.
,Matthies
м.
,м.
,
ŠULC
стр.
,Romano
F.
,Louis
A.A.
,Дойе
Дж.П.К.
,Micheletti
C.
,Rovigatti
L.
TacoxDNA: удобный веб-сервер для моделирования сложных структур ДНК, от отдельных нитей до оригами
.Дж. Вычисл. хим.
2019
;40
:2586
–2595
.46.Попплтон
Э.
,Болин
Дж.
,Маттис
М.
,Шарма
С.
,Zhang
F.
,Šulc
P.
Проектирование, оптимизация и анализ больших наноструктур ДНК и РНК посредством интерактивной визуализации, редактирования и молекулярного моделирования
.Рез. нуклеиновых кислот.
2020
;48
:E72
.47.Tian
Y.
,Wang
T.
,LIU
W.
,xin
H.L.
,LI
H.
,Ке
Ю.
,Ши
В.М.
,Gang
O.
Предписанные кластерные архитектуры наночастиц и низкоразмерные массивы, построенные с использованием каркасов оригами из октаэдрической ДНК
.Нац. нанотехнологии.
2015
;10
:637
–644
.© Автор(ы), 2021 г. Опубликовано Oxford University Press от имени Nucleic Acids Research.
Это статья в открытом доступе, распространяемая на условиях лицензии Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/), что разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы. .