ВЯЗАНЫЙ ШИК: Хризантема,МК.
ИСТОЧНИКАВТОР МК : elenanochka
ЦИТИРУЮ СЛОВА АВТОРА МК !!!
Сегодня я хочу поделиться с вами тем,как я собираю хризантему из атласных лент. Буду очень рада,если кому-нибудь пригодится моя сегодняшняя публикация.
Материалы и инструменты:
1.Атласная лента шириной 1 см двух цветов на ваше усмотрение.
2.Лента зелёного цвета шириной 5 см.
3.Проволока N°18 для стебля.
4.Флористическая проволока N°28 для листиков.
5.Тейп лента зелёного цвета.
6.Выжигатель.
7.Термопистолет.
Ленту жёлтого цвета нарезала длиной 2,5 см — 30 штук. Длина полосочек белого цвета 3,5 см. Их понадобится от 100 до 150 штук. Не пугайтесь количества,всё делается довольно-таки быстро.

Берём «ватную палочку» (извините,не знаю,как по-русски называется). С одной стороны обклеиваем кусочком ленты нужного цвета,из которого будем делать серединку нашего цветка.
И приступаем к сборке. Приклеиваем первый ряд,затем в шахматном порядке следующий… И так все тридцать лепесточков.
Вот так должно получиться. Если вы хотите сделать серединку побольше или поменьше,регулируете количеством лепестков.
Переворачиваем нашу серединку и начинаем приклеивать лепестки подлиннее. Основание лепестков приклеиваем на том же уровне,что и основания серединки.
Вот так будет выглядеть ваша работа изначально:
Таким образом приклеиваем четыре ряда лепестков. В шахматном порядке.
Вот что у нас получилось:
Лепестки следующего ряда я сдвинула на 3-4 мм к серединке. Таким образом моя хризантемка «раскрылась» к низу. На том же самом уровне приклеиваем еще два-три ряда лепестков.
Таким образом моя хризантемка «раскрылась» к низу. На том же самом уровне приклеиваем еще два-три ряда лепестков.
Обрезаем нашу «ватную палочку»,смазываем проволоку для стебля клеем и вставляем в трубочку.
Закрепляем тейп лентой,обматываем весь стебель. Выжигателем вырезаем кружочек из зелёной ленты диаметром соответствующим «донышку» цветка. Пропускаем через проволоку,предварительно сделав дырочку в серединке кружочка и приклеиваем.
Чашелистик: вырезаем выжигателем капельки и загибаем,подержав 1-2 секунды над свечой.
Приклеиваем капельки таким вот образом:
Приступаем к листикам. «Погуглила» в интернете:»лист хризантемы»,нашла понравившийся мне листочек,распечатала его,сделала шаблон из жестяной баночки. Выжигателем вырезаем листочки,им же делаем прожилочки и с изнаночной стороны вклеиваем тоненькую проволочку зелёного цвета.
 ru/node/636692 .
ru/node/636692 .Ваша хризантема готова!
Во всей красе!
С подружками!)))
[/more]МК Хризантема МК Хризантема ночник Бесплатный мастер класс – Artofit
Categories
CategoriesSelect Category360 degrees3D3d printing4K5G60 fps80s8Kabandonedabstractacrylicactoradobeadoptionaeraiaerialaerialsaerosolafricaafter effectsairplanesairportsalaskaalgorithmaliensamazonamsterdamanaloganalysisanamorphicancientancient civilizationsandroidANIMALSANIMATED GIFSANIMATED GIFSanimationanimeantarcticaapartmentsapeappapplearchaeologyARCHITECTUREarcticartart galleryarthropodsartificial intelligenceartisanastronautastronomyathensaudioaurorasaustraliaaustriaauthorsautoautomationavengersaward winningawesomeawwwbaby yodaback to the futurebalanceballoonbamboobanksybarbarbadosbarcelonabaseballbasketballbatmanbatmobilebbcbeachesbearsbeautybeesbefore and afterbehancebehaviourbehind the scenesbelgiumbernie sandersBEST OFbikingbillboardsbirdsblack and whiteblack lives matterblacksmithblendingboatsbob marleybob rossbody paintingbokehbonesbonsaibooksbosnia herzegovinabostonbowlingbrazilbreaking badbrexitbridgesbristolbronzebruce leebuddhaburning manbusinessbuster keatonbutterfliescabincakecakescalendarcaliforniacalvin and hobbescambodiacameocameracanadacandycanoecanyoncardboardcardscarl sagancartoonscarvingcarvingscarvingwcase modcastlescatcatchcatscelebrationcelebritiescell phonecemeterycentenarianceramicsceremonycgichalkchallengecharitychartschef skillschemistrychicagochinachocolatechristmaschurchciacinemagraphscity planningcity tourcityscapecityscapesclayclimate changeclose upcloudscnc millcodebreakingcodingcoffeecoinscoldplaycollaborationcollagecollectiblescolorcoloradocolorizedCOMICScomicsgcommunicationcommunitycomparisoncompetitioncompilationcompillationcompositecomputersconceptconceptualconcertconcretecondoconeptualconservationconspiracyconstructioncontestcontinuous shotcontroversialconversioncoolcoralcoronaviruscosmoscosplaycostumescountry musiccovid-19coyote and roadrunnercrabcraftcraftscraftsmanshipcreative processcrimecriminalcroatiacrochetcrocodilescross stitchcross-sectioncrowdsourcedcrystalcsscubismculturalcurrencyCURRENT EVENTScustomcyborgcyclingczech republicdalidancedarth vaderdartsdata analysisdatavizdeep learningdeepfakedefinitivedemonstrationdensitydesertDESIGNdessertdetroitdiamondsdicedigital artdigital artcdinosaursdiscoverydisneydiydnadocumentarydogsdolly partondolly parton statuedouble exposureDP Art Drawingdr seussdragon balldragonsdrawingdresdendrinkdriverdrivingdronedronesdroste effectdrumsducksdunesdysonearthearthquakeeastereaster eggsebrueco-arteconomicsedinburgheditingeducationalegypteiffel towerelderlyelectricityelectronicselephantsembroideryemojiempire state buildingengineeringenglandengravingenvironmentalerosionesaetsyeuropeeventsexperimentexplainerextremeextreme sportseyesface swapfacebookfacial recognitionfactsfailfamilyfamily guyfantasyfarmingfart jokefarthestfashionfeltfestivalfestivalefightfijiFILM/TVFILM/TVfilmsfinlandfirefireworksfirstfishflagflinstonesflipbookfloodfloor plansflowersflyingfogfontsfoodfoodcfootballforced perspectiveforestfossilfpvfractalfrancefree divingfrescofriendshipfrogsfull housefull lengthFUNNYfurniturefuturegadgetsgalaxyGALLERIESgame of thronesgamesgaminggardengardensgaudigemsgeodegeographygeologygeometrygerbilsgermanygifsglacierglassglitchglitterglowinggolfgood newsgooglegoogle earthgoogle mapsgoprogorillasgraffitgraffitigraphic designgraphsgreat big storygreat depressiongreecegreen screengreenlandGrootguerillaguitargymnasticshackhairhalloweenhamburghandmadeharpharry potterhawaiihealthhelicopterhigh-speedhikiinghikinghimalayasHISTORYhobbieshobo nickelholidayhollywoodhologramhome decorhong konghonus wagnerhorsehorseshot wheelshotelshousehousinghoustonhow its madehow tohq gallerieshtmlhuman bodyhungaryhuntinghurricanehybridhyper-realistichyperlapseibizaiceicelandikeaillusionillustrationimpressioninceptionindiaindonesiaindustrialinfographicinkinsectsinstagraminstallationinstallationainstrumentinteractiveinteriorinterior designinternetinterviewsipadiphoneiranironiron manisraelissistanbulitalyjadejamaicajapanjetsonsjewelryjokerJosh Sundquistjournalismjugglingjusticekanye westkickstarterkimmelkineticking kongkitchenkiteknittingkniveskobe bryantkurzgesagtland artlandscapelandscapeslanguagelangugelanternslargestlaserslatte artlaunchlavaleadleaveslebanonlegallegolenticularleopardletterletteringlibrarylife-sizelifehackslightlight paintinglightinglightninglighttlimestonelinkedinlinkin parklion kinglionslip syncLISTSliveloftslogoslondonlong exposurelongestlooney tuneslos angeleslouvrelovelyonlyricsmachine learningmachinesmacromad maxmaho beachmakeupmalaysiamaltamandalasmandalorianmangamanilamanufacturingmapsmarblemarblesmarketingmarsmartial artsmarvelmashupmasksmathematicsmcumedicalmedievalmelbournememesmemorabiliamemorialmentormetalmeteoritemiamimickey mantlemicro apartmentmicroscopicmicrosoftmike troutminecraftmineralsminiatureminiaturesminiautreminimalistminingmirrormmamobilemodelsmodifiedmomamona lisamonochromemontrealmonumentsmoonmoroccomosaicmostmotion capturemotivationalmotorcyclemountainsmovementmovie theatermozambiquemugshotsmumbaimummymuppetsmuppetssmuralmuseumsMUSICmysterymythologynamibianaplesNASAnat geonational geographicnational parknatureNATURE/SPACENATURE/SPACEnegative spacenerdwriternestsnetherlandsneural networknevadanew yorknew york timesnew yorkernewsnigerianightnight timenintendonoaanorwaynow and thennownessococeanoctopusodeithofficesoilolympicsomanopen sourceopenaioperaopulenceoregonorigamiosakaoscarsovergrowthoverheadowlspacific oceanpackagingpaintingpaintingspakistanpalestinepandaspandemicpanoramaspaperpaper marblingparentingparisparkourparodypastrypatternspatternswpeanutspencilpenthouseperfect timingperformanceperspectiveperuphenomenaphilippinesphosphorescentphoto seriesphotographyphotos seriesphotoshopphysicspianopicassoPICTURE OF THE DAYpikachupixarpixel artplanet earthplanetsplantsplasticpoetrypokemonpolandpolar bearpolicepolitcspoliticspollutionpompeiipoolspopeyepopulationporcelainportalportlandportraitportraitsportugalposterspotterypovpractical effectspranksprintingpro tipsproduct designprogrammingprogressprojectionproposalprotestprototypepublic spacepumpkinspuppetspuzzlepuzzle boxpuzzlegquadcopterqueenquillingquiltingquotesrabbitrainrarerawreactionreactionsreal timereclaimedrecreaterecreatevrecursivered bullredditreefsreflectionreggaerelationshipsreliefreligionrembrandtremixremote camerarenaissancerenderrepairreplicarepurposerescueresearchresinresortsrestaurantsrestorationretroreuserick and mortyrio de janeiroroadsrobberyrobotsrockrocketsrolling shutterromerooftoppingroomsroyaltyrubberrube goldbergrugsruinsrussiasalt lake citysalvador dalisamuraisan fransandsatellitesatiresaturnSCI/TECHSCI/TECHsciencescissorsscotlandsculpturesealsseasonssecretselfiesesame streetsewingsfxshadowsshakespearesharksshenzhenshipSHIRK REPORTshoesshort filmsignssilhouettesingaporeskateboardingsketchskiingskullsskyskydivingskylineskywardsleeping beautyslicedslow mo guysslow motionsmall spacessmallestsmarter every daysmartphonesmithsoniansnakessnapchatsnoopsnowsoccersocial experimentsocial mediasoftwaresolarsolar systemsonysoundsouth africasouth koreaspacespacexspainspeakersspeechesspeedspider-manspidersSPORTSspringst maartenstadiumstained glassstairsstampsstan leestarstar trailsstar trekstar warsstarsstartupstatuestatuessteampunkstep brothersstickersstill lifestockholmstonestop motionSTORIESstormstranger thingsstreamstreet artstreet photographystrongestsubculturesubvertsunrisesunsetsuper mariosuperheroessupermansurface tensionsurfingsurrealsurveysushisuspendedsvalbardswedenswitzerlandswordssydneysymmetrytable tennistaiwantapetapestrytattootaxiteamworktechtelecommunicationstempletennistensionteslatexastextiletextingthailandthanosThe Avengersthe beatlesthe simpsonsthe sunthreadtigerstiktoktilt-shifttimetime-lapsetimelapsetimelinetindertiny planettmnttokyotom and jerrytoolstoptorontotourtoy storytoystraditiontraffictrailerstrainstransformerstransparenttransportationTRAVELtreehousetreestributetrippytriptychtrompe loeiltsunamitumblrtunisiaturkeytutorialtv showstwittertypographyufoukraineUncategorizedunderwaterunescounexpectedunited statesunreal engineupcycleupliftingupscaleurban explorationus presidentutrechtvan goghvectorvehiclesvehiclestvenicevenusvespasvfxvideovideo conferencingvideo essayvintagevirtual realityvisualizationvisualizationsvoicevoiceovervolcanoesvoxwwarwashingtonwaspswatcheswaterwatercolorwaterfrontwaterspoutwaterwwavesweaponsweatherweddingweldingwhalewhaleswhat ifwikipediawinwindwinewirewiredwizard of ozwolfwoodwoodturningwoodworkwoolworld recordworld tourworld war 1writingwtfwu-tangwuhanxboxxkcdyarn bombingyodayogayoutubezoetropezombieszoomImage gallery for:
МК Хризантема МК Хризантема ночник Бесплатный мастер классМК Хризантема.
 МК Хризантема ночник. Бесплатный мастер класс — YouTube
МК Хризантема ночник. Бесплатный мастер класс — YouTube- Advertisement
- Advertisement
- Advertisement
- Advertisement
- Advertisement
 МК Хризантема ночник. Бесплатный мастер класс — YouTube
МК Хризантема ночник. Бесплатный мастер класс — YouTubeМастер-класс по валянию Хризантемы и Астры (смешаная техника).

Дорогие девочки и мальчики 🙂
Хочу поделиться с вами самым нудным простым, мастер-классом по валянию Хризантемы и Астры, который снизошел вдохновением в мою душу и реализовался через сердце и «простые движения» рук 🙂
Ну что, сваляем с помощью моего скромного толкования и фоток
ХРИЗАНТЕМА
Нам понадобится 5-6 прядей белой мериносовской шерсти для лепестков и 1 прядка желтой для серединки, пупырчатая пленка, мыльный раствор, сетка, поролоновая губка, иглы для валяния, щетка для валяния, нитка, иголка, фурнитура для брошки или заколки.
Ход работы:
1. разбираем прядки на тонененькие полосочки шерсти.
2.Каждую полосочку скручиваем и смачиваем мыльным раствором
3.Валаяем жгутики
4. После долгих, однообразных движений(я же сразу предупредила что он нудный..ну то есть простой МК 🙂 ) получаем горстку готовых мокрых лепестков (их должно быть много, порядка 50, примерно)
5. Кладем их сушить.

6. Когда лепестки высохнут берем длинную нитку, иголку и начинаем нанизывать их как зубы мамонта. Причем, возможно, у вас будет так же как и у меня — лепестки не одной длины, а разной, в этом случае нужно распределить лепестки по кучкам с примерно одинаковой длиной и начинать «ожерелье»с самых маленьких и заканчивая самыми большими.
Первыми нанизываем желтые. Еси они сильно длинные — переламываем пополам и нанизываем согнутыми.
7.Когда все лепестки нанизались, закручиваем их и начинаем прошивать основание, чтобы цветок не развалился при эксплуатации :)Нитку не отрезаем после прошивания.Ею мы потом пришьем еще лист, но если нитка мала, то тогда, делаем узел и отрезаем.
8.Начинаем делать лист. Для этого я взяла зеленый кардачес и оттенки зеленого мериноса и выложила нечто похожее на лист 🙂
9. Накрываем сеткой, намыливаем и валяем до тех пор пока волокна шерсти не превратяться в единое целое…зеленое 🙂
10.
 Прорезаем или разрываем краешки и опять замыливаем и заваливаем, вытягиваем если надо, вообщем формируем лист хризантемы 🙂
Прорезаем или разрываем краешки и опять замыливаем и заваливаем, вытягиваем если надо, вообщем формируем лист хризантемы 🙂 11. Кладем сушить.
12. Берем прядь зеленой шерсти (в тон листа) и заворачиваем и заваливаем «корешок»цветка с помощью игл.
13.Берем высохший лист и знакомим его с цветком 🙂
14.Пришиваем его иголкой и ниткой к основанию цветка. Если нитка у вас осталась и она белая, после пришивания добавляем шерсти, чтобы не было заметны белые швы на листике 🙂
15. Еще я потом вспомниа про прожилки, вы можете сначала сделать оные, а уж потом пришить лист к основанию цветка 🙂
Прожилки я делала путем выкладывания светлозеленой или белой шерстяной тонюсенькой прядки и прибивания ее иголкой к листу 🙂
16. Пришиваем фурнитуру для брошки (или заколки)к листу и вешаем бубонечку на платишко/пальтишечку или куда душа желает привесить 🙂
ну что, АСТРУ будем делать? :)))))
Нам понадобится инвентарь из МК по Хризантеме, а вот шерсть я взяла двух близких к друг другу цветов — розового и сиреневого.
1.
2.Так же как и в МК по Хризантеме, разбираем шерсть на тоненькие прядочки, скручиваем,валяем на губке,сушим, словом выполняем пункты с 1 по 5.
3. Разбираем по длине, когда лепестки высохли.(их должно быть много, штук 70 не меньше)
4. Берем самые маленькие, складываем пополам и начинаем прошивать.
5.так поступаем со всеми лепестками
6.скручиваем и прошиваем хорошо-хорошо 🙂
7.Делаем лист, примерно таким же образом как и лист хризантемы, только я выкладывала просто небольшой квадрат, без особо выраженых намеков на лист :)Для него был взят тот же зеленый кардачес и оттенки зеленого мериноса.
8. Намыливаем и сваливаем как на фотах в пункте 9 предыдущего МК.
9. Получаем нечто 🙂
10.Это нечто режем лохмотьями, оставляя цельной только середину.
11.заваливаем, путем замыливания и вытягивания лохмотьев :)Сушим.
12 получили:
13. Прибиваем полученые лохмотья к прошитому корешку астры.
 Где необходимо добавляем шерсти в тон.
Где необходимо добавляем шерсти в тон. 14. получам:
15. пришиваем фурнитуру для брошки(заколки)
16. украшаем себя любимую …или свою любимую :))))
ну что…сваляем ГЕОРГИН? :))))
Tags: а я делаю так!, аксессуары 2011, мокрое валяние 2011, сухое валяние 2011
Букет хризантем из бумаги – Уроки Рукоделия
Поделитесь ссылкой на эту статью с друзьями в социальных сетях.
Создание цветов из бумаги занятие довольно-таки увлекательное. К тому же сделать бумажные цветочки не так уж сложно, главное удачно подобрать технику исполнения и правильно выполнить все указанные действия. Сегодня существует огромное количество интересных способов их создания, поэтому даже начинающая рукодельница найдет ту технику, которая ей под силу. Готовыми цветами из бумаги вы сможете оригинально украсить свое жилище или даже вручить их в качестве подарка. В данном мастер-классе мы вам покажем, как самостоятельно сделать хризантему из гофрированной бумаги.
Приготовьте необходимые материалы:
- гофрированная бумага желаемого оттенка для создания лепестков;
- зелёная гофро-бумага;
- ножницы;
- клеевой пистолет;
- простой карандаш;
- проволока;
- линейка.
Шаг 1
Начнем работу с создания стебля хризантемы. Приготовим тонкую ленточку гофрированной бумаги зелёного оттенка, отрезок проволоки длиной, приблизительно, 30 см и термоклей.
Прежде чем мы начнем обматывать бумажной полоской проволоку, разглаживаем все её складки. Для этого просто растяните ленту в разные стороны и прогладьте её кончиками пальцев.
Далее наносим на начало ленты капельку клея, присоединяем её к кончику проволоки и начинаем обматывать ею весь отрезок.
Конец ленты фиксируем клеем.
Шаг 2
Для создания лепестков отрежьте от рулона полосу шириной 5см.
Далее отмечаем на ленте 4,5 см. и разрезаем ленту до поставленной точки.
Ширина такой полосочки должна быть не более 1 см.
Продолжаем разрезать таким же образом оставшийся кусочек бумажной ленты.
Шаг 3
Теперь полоскам нужно придать форму лепестков. Верхнюю часть полоски перекручиваем, затем сгибаем ее и складываем ко второй части полосы.
После растягиваем ее кончик, чтобы получилось небольшое углубление.
Верхнюю часть полоски приклеиваем к нижней.
Продолжаем создавать вот такие лепесточки из остальных полосок.
Для создания одного бутона хризантемы нам понадобится три полоски с лепестками-заготовками.
Шаг 4
Далее присоединяем все ленточки с лепестками. Наносим клей на внутреннюю часть первого лепесточка и на кончик стебля, присоединяем к верхней части стебля и начинаем накручивать её.
Постепенно бутон будет приобретать красивую форму.
Конец полосок фиксируем клеем.
Шаг 5
Чашелистики вырезаем из зелёной гофро-бумаги.
Растягиваем заготовки, чтобы они получились вот такой формы.
Затем приклеиваем их к основе соцветия, скрывая внутреннюю часть бутона. А основание чашелистиков обматываем зелёной гофро-полоской.
Кончики чашелистиков можно немного подкрутить, придавая естественную форму.
Шаг 6
Теперь можно вырезать из зелёной гофро-бумаги листья хризантемы вот такой формы.
Присоединяем их к веточке с помощью термоклея.
Вот и все, реалистичные хризантемы готовы!
Если вы любите крупные цветы с объёмными лепестками, то сделайте хризантемы.
Такие цветы будут хорошо смотреться в любом интерьере в виде обычного букетика или композиции.
Красота бумажных цветов поражает своей схожестью с живыми, они завораживают собой и восхищают взгляды. Из гофрированной бумаги можно сделать не только хризантемы, но и другие цветы.
Мастер-класс от Лёлика.
Комплект постельного белья Chrysanthemum MkCyliym — DalabShop
Вы очень занятый человек и у вас мало времени на отдых. Действительно, первое, что хочется сделать после работы, это лечь на удобную кровать.
Действительно, первое, что хочется сделать после работы, это лечь на удобную кровать.
Вам не составит труда найти комплект постельного белья. При этом вы можете выбирать из множества моделей с разной стоимостью.
Попробуйте комплект постельного белья Chrysanthemum Mk Cyliym, если хотите вернуться в детство. Или, если у вас есть дети, которые любят научно-фантастическую анимацию, подарите им этот комплект постельного белья.
Что делает этот уникальный дизайн привлекательным? Давайте разберемся!
Материал
Как и большинство товаров, представленных на рынке, комплект постельного белья изготовлен из полиэфирной ткани. Сегодня это самая распространенная ткань на рынке.
Отличительной чертой этого материала является то, что он меньше мнется, пыленепроницаемый, водостойкий и легко моется, что экономит время на стирку или разглаживание простыней после каждого сна.
При этом продукт также безопасен для кожи всех членов семьи.В частности, они безопасны для детей и не вызывают раздражения.
Одной из особенностей этой ткани, которую любит каждый ребенок, является ее способность легко окрашиваться. Поэтому этот материал четко покажет цвета и характеристики персонажа при печати. При этом изображение, напечатанное на ткани, не выцветает при стирке с другой одеждой или тканями.
Качество
Несмотря на то, что комплект постельного белья Chrysanthemum Mk из химических волокон изготовлен из химических волокон, он очень мягкий и не вызывает раздражения или аллергии даже на самой нежной коже.
Наволочки также помогают бороться с прыщами на лице из-за их незначительного запыления. Это то преимущество, на которое многие обращают внимание при выборе постельного гарнитура. Тем не менее, вы также должны регулярно стирать постельный комплект, чтобы предотвратить максимальное проникновение грязи.
Это выбор, который стоит попробовать с точки зрения материала и ощущения продукта. Многие люди, купившие этот постельный комплект, хорошо отзываются о его мягкости и выдающихся достоинствах
.Компоненты и размер
Chrysanthemum Mk Комплект постельного белья Cyliym включает в себя наволочку и пододеяльник.
Наволочки стандартного размера для членов семьи, размером 48 см x 73 см, подходят для обычной подушки в спальне.
В отличие от наволочек, пододеяльники бывают разных размеров. Вы можете выбрать пододеяльник в зависимости от типа вашей кровати. Пододеяльник имеет односпальную кровать, двуспальную кровать, размер королевы и даже размер короля.
Пододеяльник шириной 145 см и длиной 210 см для односпальной кровати. Точно так же двуспальная кровать имеет размеры 180 см x 210 см для кроватей размера «queen-size» и «king-size» с размерами 210 см x 210 см и 220 см x 240 см соответственно.
Выбор подушек и одеял рядом с наволочками необходим для того, чтобы ваша кровать стала удобной, особенно изделия из натуральных перьев.
Цвет
Вдохновленный комплектом постельного белья Chrysanthemum Mk Cyliym, комплект постельного белья станет изображением, наиболее реалистично описывающим характеристики и историю персонажа.
Благодаря полиэфирной ткани комплект постельного белья не только легко печатается на поверхности, но и помогает комплекту сохранять свой цвет и качество в течение длительного времени.
Постельный комплект можно стирать в стиральной машине, не беспокоясь о сминании или выцветании других изделий. Эта особенность заставляет многих мам доверять и выбирать при покупке комплекта матраса для своего ребенка.
Некоторые часто задаваемые вопросы
Как осуществляется доставка продукта?
Вы можете найти товары на нашем веб-сайте электронной коммерции со многими формами оплаты.
Вы получите товар в течение 20 рабочих дней. При этом вам также будет предоставлен номер заказа для отслеживания коносамента.Если продукт отправляется за границу, уплата налогов и сборов будет на вас.
Как стирать постельный комплект?
Изделие изготовлено из полиэстера, поэтому быстро не выгорает и очень прочно в обычных условиях. Так что стирать постельный комплект можно в стиральной машине или вручную.
Не следует использовать отбеливатель с высоким уровнем pH, так как постельный комплект легко пожелтеет или потускнеет.
В то же время, вы должны использовать стиральный порошок и смягчитель, которые ароматизированы и безопасны для кожи, чтобы продукт был приятным.Это не только поможет сохранить кровать чистой и без плесени, но также поможет вашему ребенку комфортно заснуть и расслабиться.
Заключение
В приведенной выше статье представлены все характеристики комплекта постельного белья Chrysanthemum Mk Ciliym. Это не просто продукт для спальни, он также может помочь вашему ребенку стать более творческим и полюбить науку.
Это не просто продукт для спальни, он также может помочь вашему ребенку стать более творческим и полюбить науку.
Не медлите! Дайте отличный продукт для сна вашего ребенка.
img
: Удалено
К сожалению, ID элемента: img
не найден.
19C China 2 Ваза для разливов из цельного серебра Repousse Цветы и бамбук с маркировкой 九江涂茂興
asia-antika
ПРЕДЛОЖЕНИЕ $500
Китайская фарфоровая банка Famille Rose Noire 19C, горшок, ваза, цветок сливы, цветущая птица
asia-antika
ПРЕДЛОЖЕНИЕ $2500
Английская тарелка Worcester Kakiemon с мягкой пастой 18-го века, дольчатая перепелиная тарелка
asia-antika
ПРЕДЛОЖЕНИЕ $950
Vintage Oscar Bucher California Crafts Flame Glaze Керамическая китайская чаша
asia-antika
ПРЕДЛОЖЕНИЕ $300
Китайское позолоченное серебро 1930-х годов Нефрит Жадеит Коралл Резные бусины Заколка с маркировкой
asia-antika
ПРЕДЛОЖЕНИЕ $150
Статуэтка японской школы Токио 1900-х годов, бронзовая фигурка мальчика, лови птицу, фигурка Окимон Sg
asia-antika
ПРЕДЛОЖЕНИЕ $1500
Индия Индийский индуистский джуйпар Деревянная подставка из цельного серебра с эмалью Павлин
asia-antika
ПРЕДЛОЖЕНИЕ $950
19C Рог японского оленя Резная трость Трость Ручка для зонтика
asia-antika
ПРЕДЛОЖЕНИЕ $750
Китайский нефрит Жадеит Резная табличка Журавль Серебряная эмаль Филигранный браслет Mk
asia-antika
ПРЕДЛОЖЕНИЕ 1000 $
2 Винтажная маленькая ваза с японской розовой перегородчатой эмалью Shippo Roses Flower
asia-antika
ПРЕДЛОЖЕНИЕ $250
Японская студия Peach Bloom Flambe Glaze Porcelain Scholar Censer Inoue Ryosai
asia-antika
ПРЕДЛОЖЕНИЕ $500
Ожерелье из китайского позолоченного серебра 1930-х годов из лососевого коралла с резным кулоном и жемчугом
asia-antika
ПРЕДЛОЖЕНИЕ $450
Китайский золотой бриллиант 14 карат, шпинат, нефрит, резная брошь, булавка, маркировка
asia-antika
ПРЕДЛОЖЕНИЕ $500
Яичный желток, ириска, резная янтарная брошь, 19 грамм
asia-antika
ПРЕДЛОЖЕНИЕ $200
1920-е годы Ар-деко Сумочка из микростеклянных бусин Кошелек Цветы Бабочка Шнурок
asia-antika
ПРЕДЛОЖЕНИЕ $250
Японский фарфор Кутани Каннон Куан Инь Кван Инь Статуэтка Будды
asia-antika
ПРЕДЛОЖЕНИЕ $250
Китайский набор 1940-х годов, 8 серебряных агатовых резных сердоликов, чайная ложка Mk
asia-antika
ПРЕДЛОЖЕНИЕ $500
Японский смешанный металл 1930-х годов, стиль Komai, дамасская пряжка для ремня, маркировка Phoenix
asia-antika
ПРЕДЛОЖЕНИЕ $350
Шелковый вышитый веер из китайской бычьей буйволиной кости 1930-х годов
asia-antika
ПРЕДЛОЖЕНИЕ $300
Фарфоровое колье с ручной росписью и хризантемами из китайских фамильных роз
asia-antika
ПРЕДЛОЖЕНИЕ $200
Memorial Sloan Kettering Cancer Center
Отказ от ответственности
Этот веб-сайт — Информация о травах, растительных препаратах и других продуктах — предназначен только для общей медицинской информации. Этот веб-сайт не может использоваться в качестве замены медицинской консультации, диагностики или лечения какого-либо состояния или проблемы со здоровьем. Пользователи этого веб-сайта не должны полагаться на информацию, предоставленную на этом веб-сайте, для решения своих проблем со здоровьем. Любые вопросы, касающиеся вашего собственного здоровья, должны быть адресованы вашему лечащему врачу или другому поставщику медицинских услуг.
Этот веб-сайт не может использоваться в качестве замены медицинской консультации, диагностики или лечения какого-либо состояния или проблемы со здоровьем. Пользователи этого веб-сайта не должны полагаться на информацию, предоставленную на этом веб-сайте, для решения своих проблем со здоровьем. Любые вопросы, касающиеся вашего собственного здоровья, должны быть адресованы вашему лечащему врачу или другому поставщику медицинских услуг.
Memorial Sloan Kettering Cancer Center не дает никаких гарантий и не делает явных или подразумеваемых заявлений относительно точности, полноты, своевременности, сравнительного или противоречивого характера или полезности любой информации, содержащейся или упоминаемой на этом веб-сайте.Memorial Sloan Kettering не принимает на себя никакого риска в связи с использованием вами данного веб-сайта или содержащейся на нем информации. Информация о здоровье часто меняется, поэтому информация, содержащаяся на этом веб-сайте, может быть устаревшей, неполной или неправильной. Заявления, сделанные о продуктах, не были оценены Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов. Использование данного веб-сайта не создает выраженных или подразумеваемых отношений между врачом и пациентом.
Заявления, сделанные о продуктах, не были оценены Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов. Использование данного веб-сайта не создает выраженных или подразумеваемых отношений между врачом и пациентом.
Memorial Sloan Kettering не записывает конкретную информацию о пользователях веб-сайта и не связывается с пользователями этого веб-сайта.Настоящим вам рекомендуется проконсультироваться с врачом или другим профессиональным поставщиком медицинских услуг, прежде чем принимать какие-либо решения или предпринимать какие-либо действия или не предпринимать никаких действий, связанных с какой-либо медицинской проблемой или проблемой, которая может возникнуть у вас в любое время, сейчас или в будущем. будущее. Используя этот веб-сайт, вы соглашаетесь с тем, что ни Memorial Sloan Kettering, ни какая-либо другая сторона не несут и не будут нести иной ответственности за любое принятое решение или любое предпринятое действие или любое действие, не предпринятое в связи с использованием вами какой-либо информации, представленной на этом веб-сайте.
Для пациентов и лиц, осуществляющих уход
Сообщите своим поставщикам медицинских услуг о любых пищевых добавках, которые вы принимаете, таких как травы, витамины, минералы и натуральные или домашние средства. Это поможет им управлять вашим уходом и обеспечить вашу безопасность.
Хризантема не лечит и не предотвращает рак.
Хризантема — цветущее растение из семейства подсолнечных. Он использовался в традиционной медицине на протяжении веков, но исследований было проведено мало.Лабораторные исследования показывают, что это может быть полезно для лечения заболеваний костей и диабета. Некоторые исследования показывают, что экстракты хризантемы могут убивать раковые клетки в лаборатории, но неизвестно, проявляется ли этот эффект в организме человека.
Пациентам, получающим препараты для подавления иммунной системы, следует избегать этого растения, так как оно может усилить побочные эффекты, связанные с этими препаратами.
- Для лечения стенокардии
Хризантема используется для лечения стенокардии в традиционной китайской медицине, но исследования не проводились. - Для профилактики и лечения простуды
Хотя хризантема используется для лечения простуды в традиционной китайской медицине, ее применение на людях не изучалось. - Для снижения температуры
Хризантема используется в качестве жаропонижающего средства в традиционной китайской медицине, но данные о людях отсутствуют. - Для снижения высокого кровяного давления
Хотя хризантема используется для лечения высокого кровяного давления в традиционной китайской медицине, клинических исследований не проводилось. - Для уменьшения воспаления
Лабораторные исследования свидетельствуют о различных свойствах хризантемы, включая противовоспалительное действие, но исследования на людях отсутствуют.

- Вы принимаете препаратов для подавления иммунной системы: У реципиента почки, который пил чай, содержащий хризантему, были обнаружены токсичные уровни этих препаратов в крови, и лабораторный анализ подтвердил, что хризантема, вероятно, была способствующим фактором.
- Вы принимаете Cytochrome P450 3A4 или препараты субстрата P-гликопротеина: Chrysanthemum может изменить их действие.
- У вас аллергия на амброзию .

- Покраснение, отек и зуд кожи
- Аллергическая реакция
- Повышенная чувствительность к солнечному свету и вероятность получения солнечных ожогов
Отчеты о случаях заболевания
Кожная сыпь: При профессиональном контакте с хризантемой.
Токсические уровни иммунодепрессантов в крови: У реципиента почечного трансплантата, который произошел после употребления чая «24 вкуса». Было установлено, что одно из ключевых соединений чая, хризантема, ингибирует фермент, который метаболизирует эти лекарства.
Хризантема – советы по выращиванию и уходу для мам
Хризантемы (мамочки) – одни из самых популярных осенних садовых цветов. Большинство маточных растений легко выращивать, их основными потребностями являются полное солнце, богатая почва, хороший дренаж и хорошая циркуляция воздуха.Существуют сотни сортов, доступных в различных формах и размерах, которые могут обеспечить цветение с конца лета до осени. Хризантемы символизируют разные вещи в разных странах: жизнь и возрождение в Азии, симпатию в Европе, уважение и почет в Америке.
На этой странице: Основы | Посадка и уход | Идеи дизайна | Фотографии | Часто задаваемые вопросы
ОСНОВЫ
Розовая садовая мамочка Paradiso. Фото: Проверенные победители.
Зоны:
5-9 (некоторые разновидности для зоны 4).
Высота/ширина:
Разновидности 1–3 фута/1–2 фута
Цветок Цвет:
Цветки хризантемы распускаются в различных оттенках белого, желтого, оранжевого, лавандового, пурпурного или красного; а также двухцветные цветы.
Воздействие:
Предпочитают полное солнечное освещение в течение вегетационного периода. Недостаток солнечного света приведет к более слабому растению, которое производит меньше цветочных головок. Тем не менее, цветение будет длиться дольше, если его переместить в более затененное место после того, как появятся цветочные почки.
Почва:
Мамы предпочитают плодородную, хорошо дренированную почву. Есть хорошее эмпирическое правило: если почва хороша для овощей, она хороша и для мам.
Время цветения:
Сентябрь до заморозков. Известные как растения короткого дня, цветение вызвано более короткими днями в конце лета и начале осени. Цветение также можно форсировать в теплице с регулируемым освещением.
КАК ПОСАДИТЬ МАМ И УХОД ЗА МАМОЙ
Когда сажать:
Для использования в качестве многолетника сажайте ранней весной или по крайней мере за 6 недель до осенних морозов.Посадка хризантем весной даст им больше шансов пережить следующую зиму. Если вы используете их в качестве однолетних растений осеннего цвета, сажайте их во время цветения в конце лета или в начале осени.
Если вы используете их в качестве однолетних растений осеннего цвета, сажайте их во время цветения в конце лета или в начале осени.
Вода:
Мамы требуют частого полива из-за их неглубокой корневой системы, особенно в жару. Слой мульчи летом поможет сохранить воду и сохранить почву влажной и прохладной.
Обрезка:
Прищипывайте примерно 1 дюйм от кончиков ветвей два-три раза в течение вегетационного периода, чтобы стимулировать ветвление и сделать растение более крепким.Раннецветущие, которые цветут в середине сентября, следует прищипывать не позднее середины июня. Октябрьские цветки можно прищипывать до середины июля, руководствуясь эмпирическим правилом: не прищипывать ближе, чем за 3 месяца до цветения.
Размножение:
При выращивании как многолетники их можно делить весной каждые два-три года. Выкопайте растение, когда начнет появляться новый рост, удалите отмирающий центр и повторно посадите новые побеги снаружи растения. Их также можно вырастить из черенков, взятых весной. Срежьте чуть ниже листового узла и укорените в стерильной почве. Новые растения следует ежедневно поливать и держать на солнечном подоконнике, пока они не приживутся.
Срежьте чуть ниже листового узла и укорените в стерильной почве. Новые растения следует ежедневно поливать и держать на солнечном подоконнике, пока они не приживутся.
Удобрение:
Мамы не любят много кормить, поэтому лучше несколько раз вносить разбавленное удобрение до образования бутонов. Состав удобрения 5-10-5 окажет наибольшее влияние на цветение и общий рост.
Болезни и вредители:
Некоторыми болезнями, которые могут поразить их, являются пятнистость листьев, мучнистая роса и вирусные заболевания, такие как мозаика или кашель.Избегайте перенаселенности и слишком тенистых мест, где влага остается на листьях и создает среду обитания для болезней. Вредителями могут быть тли, гусеницы, цикадки, минеры, жуки и паутинные клещи.
ИДЕИ ДИЗАЙНА
Создайте осенний сказочный сад  5em;text-align:center;font:48px/1.5 sans-serif;color:white;text-shadow:0 0 0.5em black}</style><a href=https://www.youtube.com/embed/AJxCQhbW97c?autoplay=1><img src=https://i.ytimg.com/vi/AJxCQhbW97c/maxresdefault.jpg alt=’Fall Fairy Garden’><span>▶</span></a>» frameborder=»0″ allow=»accelerometer; autoplay; encrypted-media; gyroscope; picture-in-picture» allowfullscreen=»»/>
5em;text-align:center;font:48px/1.5 sans-serif;color:white;text-shadow:0 0 0.5em black}</style><a href=https://www.youtube.com/embed/AJxCQhbW97c?autoplay=1><img src=https://i.ytimg.com/vi/AJxCQhbW97c/maxresdefault.jpg alt=’Fall Fairy Garden’><span>▶</span></a>» frameborder=»0″ allow=»accelerometer; autoplay; encrypted-media; gyroscope; picture-in-picture» allowfullscreen=»»/>
Хризантемы отлично подходят для выращивания прямо в саду или в контейнерах, вот несколько идей:
- Идеально подходит для выращивания в контейнерах и корзинах благодаря неглубокому укоренению.
- Используйте осенью как однолетник для замены цветущих летом однолетников.
- Имея множество доступных цветовых вариантов, мамы могут расставить координирующие или контрастные цветовые акценты как в помещении, так и на открытом воздухе.
- Они относительно недороги, что делает их отличным выбором для больших групп или повторения по всей территории.
- Растите рядом с другими цветущими осенью растениями, такими как эхинацея, очиток, астра, голубая борода и анютины глазки.

САДОВАЯ МАМА ПРОТИВ.МАМА-ФЛОРИСТ:
Разница между мамами-садоводами и цветоводами сводится к их выносливости. Садовые мамы — это, как правило, сорта, которые вы бы посадили на открытом воздухе в своем саду, и они выносливы в своих определенных зонах. Мамы-флористы используются исключительно для выращивания комнатных растений в горшках и не подходят для пересадки на улицу. Убедитесь, что вы покупаете правильный тип для вашего предполагаемого использования и местоположения.
ФОТОГРАФИИ
Проведите пальцем для просмотра слайдов
Фото: Проверенные победители.
Фото: Проверенные победители.
фламинго розовый ананас по договорной цене в Proven Winners
C. grandiflorum
Зоны: 7-9
Время цветения: С конца лета до осени
Мамочка уникального цвета, устойчивая как к жаре, так и к морозу. Более высокие температуры приводят к более кремовому цвету; более низкие температуры, более розовый.
Более высокие температуры приводят к более кремовому цвету; более низкие температуры, более розовый.
Фото: Проверенные победители.
розовый paradiso по договорной цене в Proven Winners
C.крупноцветковый
Зоны: 7-9
Время цветения: С конца лета до осени
Добавьте темно-розовый оттенок своим осенним выставкам. Вырастает от 12 до 20 дюймов в высоту и до 30 дюймов в ширину. Также доступны в желтом и бронзовом цвете в этой серии.
Фото: Проверенные победители.
Stacy™ Dazzling Orange — Купить у Proven Winners
C. grandiflorum
Зоны: 7-9, обычно выращивается как однолетник
Время цветения: С конца лета до поздней осени
Сочетая два классических цвета мамы, оранжевый и желтый, этот сорт устойчив к морозам, поэтому ваш сад будет красочным в течение недели в осень. Привлекающее бабочек и простое в выращивании, добавьте это в свой список «must have»!
Привлекающее бабочек и простое в выращивании, добавьте это в свой список «must have»!
Фото: Проверенные победители.
желтый сандэнс по договорной цене в Proven Winners
C. grandiflorum
Зоны: 7-9, обычно выращивается как однолетник
Время цветения: С конца лета до поздней осени
Добавьте эту веселую желтую маму в свои контейнеры, чтобы пригласить колибри и бабочек в осенний сад. Подарите себе немного счастья, потому что из этой мумии получаются отличные срезанные цветы!
Фото: Проверенные победители.
красный моргана по договорной цене в Proven Winners
C. grandiflorum
Зоны: 7-9, обычно выращивается как однолетник
Время цветения: С конца лета до поздней осени
Ростом 12–20 дюймов, эта темно-красная мама отлично подходит для контейнеров, пейзажей и на срезку. цветок для создания классического осеннего интереса и привлечения бабочек.
цветок для создания классического осеннего интереса и привлечения бабочек.
Фото: Bluestone Perennials.
Хризантема «Возвращение домой»
Зоны: 5-9
Время цветения: Осень
Форма цветка: Футбол
‘Homecoming’ — потрясающая мама с пышными крупными цветками; идеально подходит для резки.
Фото: Bluestone Perennials.
Хризантема Спички
Зоны: 5-9
Время цветения: Осень
Форма цветка: Ложка/перо
Яркий двухцветный цветок, который иногда распускается летом, а затем снова осенью.
Фото: Martin Hughes-Jones / Alamy Stock Photo.
Хризантема «Бронзовый Элеганс»
Зоны: 6-9
Время цветения: Осень
Форма цветка: Махровый
У этой старомодной мамы полностью махровые цветки идеальной осенней окраски.
Фото: B Christopher / Alamy Stock Photo.
Хризантема «Королева Камбоджи»
Зоны: 4-9
Время цветения: Сентябрь до заморозков
Форма цветка: Маргаритка
Спорт сорта ‘Шеффилд Пинк’ с более глубоким розовым оттенком. Выращивается в Ботаническом саду Прибрежного штата Мэн, Ботаническом саду Норфолка и Ботаническом саду США.
Фото: Garden World Images Ltd / Alamy Stock Photo.
Хризантема «Рубиновый холм»
Зоны: 5-9
Время цветения: август-октябрь
Форма цветка: Подушечное
Сорт по зимостойкости в Миннесоте, эта мама классифицируется как декоративная, что означает, что это компактное, округлое растение с формальный цветок с большим количеством лепестков. Популярен для бордюров и коттеджных садов.
Фото: Garden World Images Ltd / Alamy Stock Photo.
Хризантема ‘Желтая звездочка’
Зоны: 5-9
Время цветения: Сентябрь до заморозков
Форма цветка: Ложка
Эта мама выше, достигает до 2 футов в высоту.
Фото: Garden World Images Ltd / Alamy Stock Photo.
Хризантема ‘Карие глаза’
Зоны: 5-9
Время цветения: Сентябрь до заморозков
Форма цветка: Пуговица
Образует низкий холмик с жесткими цветками.
Фото: Rex May / Alamy Stock Photo.
Хризантема «Император Китая»
Зоны: 4-9
Время цветения: август-октябрь
Форма цветка: Изогнутая
Листья этой мумии с возрастом становятся темно-красными, вызывая дополнительный интерес к падению.
ЧАСТО ЗАДАВАЕМЫЕ ВОПРОСЫ
Мамы однолетние или многолетние?
Несмотря на то, что самые распространенные мамы выращивают как осенние однолетники, существуют сорта, которые также можно выращивать как многолетники в некоторых регионах с небольшим уходом за ними на зимовку. В более холодных зонах оставьте верхние побеги на месте и добавьте рыхлую мульчу, такую как солома или вечнозеленые ветки вокруг них для защиты, ожидая обрезки весной после появления новых побегов. В более теплом зимнем климате их можно обрезать до 6 дюймов в высоту после цветения.Обратитесь в местный садовый центр, чтобы узнать, какие сорта лучше всего подходят для вашего региона.
В более холодных зонах оставьте верхние побеги на месте и добавьте рыхлую мульчу, такую как солома или вечнозеленые ветки вокруг них для защиты, ожидая обрезки весной после появления новых побегов. В более теплом зимнем климате их можно обрезать до 6 дюймов в высоту после цветения.Обратитесь в местный садовый центр, чтобы узнать, какие сорта лучше всего подходят для вашего региона.
А как насчет мам в горшках, которые продаются в супермаркетах и крупных магазинах?
Эти доступные по цене растения появляются в магазинах осенью и имеют аккуратную горку маленьких цветов. Эти растения, которые иногда называют садовыми мамами или бельгийскими мамами, обычно выращивают как однолетники. Они популярны для использования в осенней экспозиции контейнеров. Большинство садоводов сообщают, что они не так выносливы, как описанные выше мамы, и редко перезимовывают.
Ядовиты ли мамы для кошек, собак или других животных?
Мамы содержат вещества, токсичные для кошек, собак и лошадей при проглатывании, поэтому обязательно имейте это в виду при выборе места, которое могут исследовать любопытные домашние животные. Узнайте больше о распространенных ядовитых растениях для собак и кошек.
Узнайте больше о распространенных ядовитых растениях для собак и кошек.
Устойчивы ли мамы оленей и кроликов?
Олени и кролики, как правило, избегают растений с ароматными листьями и ворсистой текстурой, которые являются атрибутами мам, что делает их довольно устойчивыми.
Связанный:
Встреча выпускников: техасская традиция
Астры
Выращивание герберы
Полногеномная сборка De novo Chrysanthemum makinoi, ключевой дикой хризантемы | Гены G3|Геномы|Генетика
Аннотация
Хризантема входит в десятку лучших срезанных, горшечных и многолетних садовых цветов в мире. Несмотря на это, на сегодняшний день секвенированы и собраны только геномы двух диких диплоидных хризантем.Здесь мы представляем наиболее полную и непрерывную сборку хризантемы de novo , опубликованную до сих пор, а также соответствующую аннотацию ab initio . Считается, что культурные гексаплоидные сорта произошли от гибрида диких хризантем, среди которых упоминалась диплоидная Chrysanthemum makinoi . Используя комбинацию длинных ридов Oxford Nanopore, длинных ридов Pacific Biosciences, коротких ридов Illumina, последовательностей «ласточкин хвост» и генетической карты, мы собрали 3.1 ГБ своей последовательности на девять псевдохромосом с N50 330 МБ и полной оценкой BUSCO 92,1%. Наш конвейер аннотаций ab initio предсказал 95 074 гена и пометил 80,0% генома как повторяющиеся. Эта сборка генома C. makinoi обеспечивает важный шаг вперед в понимании генома, эволюции и истории хризантемы.
Используя комбинацию длинных ридов Oxford Nanopore, длинных ридов Pacific Biosciences, коротких ридов Illumina, последовательностей «ласточкин хвост» и генетической карты, мы собрали 3.1 ГБ своей последовательности на девять псевдохромосом с N50 330 МБ и полной оценкой BUSCO 92,1%. Наш конвейер аннотаций ab initio предсказал 95 074 гена и пометил 80,0% генома как повторяющиеся. Эта сборка генома C. makinoi обеспечивает важный шаг вперед в понимании генома, эволюции и истории хризантемы.
Введение
В качестве одной из наиболее экономически важных декоративных культур (Anderson 2007) много времени было потрачено на изучение Chrysanthemum morifolium Ramat.и родственные ему разновидности и виды. Одним из ключевых факторов его успеха в качестве декоративной культуры является разнообразие цветов лепестков и форм цветков (Song et al. 2018), хотя лежащие в основе геномные и молекулярные основы признаков формы все еще плохо изучены. Отчасти это связано с тем, что это гексаплоид с полисомным наследованием (van Geest et al. 2017b).
Отчасти это связано с тем, что это гексаплоид с полисомным наследованием (van Geest et al. 2017b).
Чтобы начать понимать гексаплоид, такой как C. morifolium Ramat.и его черты, мы должны сначала взглянуть на весь род и исследовать происхождение растения. Род Chrysanthemum состоит из видов с основным числом хромосом девять, но с переменным уровнем плоидности, от диплоидного до декаплоидного (Wang et al. 2014). Этот род произрастает в Евразии и северных частях Северной Америки и состоит из 40 различных видов (Liu et al. 2012; Liu 2020). Более 10 первоначально были идентифицированы как потенциальный исходный материал для одомашненного C.морифолиум Рамат. (HEMSley 1889; Stapf 1933; Dowrick 1952; Ackerson 1967), в том числе хризантема MakinoI (SYN. D. Makinoi ), Chrysanthemum RISTIM (SYN. D. INDINGUM ), Chrysanthemum lavandulifolium (син. D. lavandulifolium ) и Chrysanthemum zawadskii (син. D. zawadskii ), преимущественно в гексаплоидной форме. Гексаплоид Chrysanthemum vestitum и тетраплоид C.indicum позже снова были предложены в качестве основных доноров на основании сравнительной морфологии, цитологии, межвидовой гибридизации и молекулярной систематики (Ma et al. 2016). Диплоиды, такие как хризантема Nankingense , C. Lavandulifoium и C. Zawadskii также неоднократно определены как возможные участники (Dai et al. 2005; Liu et al. 2012; MA et и др. 2016). На сегодняшний день никто не придумал окончательной модели для C.морифолиум Рамат.
D. lavandulifolium ) и Chrysanthemum zawadskii (син. D. zawadskii ), преимущественно в гексаплоидной форме. Гексаплоид Chrysanthemum vestitum и тетраплоид C.indicum позже снова были предложены в качестве основных доноров на основании сравнительной морфологии, цитологии, межвидовой гибридизации и молекулярной систематики (Ma et al. 2016). Диплоиды, такие как хризантема Nankingense , C. Lavandulifoium и C. Zawadskii также неоднократно определены как возможные участники (Dai et al. 2005; Liu et al. 2012; MA et и др. 2016). На сегодняшний день никто не придумал окончательной модели для C.морифолиум Рамат.
Хризантема makinoi — диплоидный эндемик Японии. Хотя в прошлом проводились исследования этого диплоидного вида (Tanaka 1960; Tanaka and Shimotomai 1968), никто не пытался собрать его геном. Фактически, на сегодняшний день из 40 видов хризантем только C hrysanthemum seticuspe (Hirakawa et al. 2019) и C. nankingense (Song et al. 2018) имеют полногеномные сборки.Сборка C. seticuspe была создана с использованием только короткого секвенирования и имела общую длину 2,722 Гб, 354 212 контигов, N50 44 741 п.н. и оценку BUSCO 88,8% (Hirakawa et al. 2019). , в то время как C. nankingense был собран с использованием как длинных, так и коротких чтений общей длиной 2,527 Гб, с 24 051 контигом, N50 130 678 п.н. и оценкой BUSCO 92,7% (Song et al. 2018). Создание более непрерывной сборки этих диплоидов было затруднено, поскольку геномы хризантем очень повторяющиеся и гетерозиготные (Won et al. 2018а; Нгуен и др. 2020).
Фактически, на сегодняшний день из 40 видов хризантем только C hrysanthemum seticuspe (Hirakawa et al. 2019) и C. nankingense (Song et al. 2018) имеют полногеномные сборки.Сборка C. seticuspe была создана с использованием только короткого секвенирования и имела общую длину 2,722 Гб, 354 212 контигов, N50 44 741 п.н. и оценку BUSCO 88,8% (Hirakawa et al. 2019). , в то время как C. nankingense был собран с использованием как длинных, так и коротких чтений общей длиной 2,527 Гб, с 24 051 контигом, N50 130 678 п.н. и оценкой BUSCO 92,7% (Song et al. 2018). Создание более непрерывной сборки этих диплоидов было затруднено, поскольку геномы хризантем очень повторяющиеся и гетерозиготные (Won et al. 2018а; Нгуен и др. 2020).
Данные длительного чтения помогают разрешить повторяющиеся последовательности и позволяют собирать больше смежных контигов (van Dijk et al. 2018), поэтому мы использовали подход, сочетающий методы длительного чтения, короткого чтения и лигирования по близости. для создания действительно надежной сборки. Эта сборка вместе с соответствующими сборками органелл и транскриптомом не только расширит наше понимание диплоида C. makinoi , но и поможет пролить свет на сложную историю полиплоидизации, которая привела к C.морифолиум Рамат. путем обеспечения надежной геномной основы для расширения.
2018), поэтому мы использовали подход, сочетающий методы длительного чтения, короткого чтения и лигирования по близости. для создания действительно надежной сборки. Эта сборка вместе с соответствующими сборками органелл и транскриптомом не только расширит наше понимание диплоида C. makinoi , но и поможет пролить свет на сложную историю полиплоидизации, которая привела к C.морифолиум Рамат. путем обеспечения надежной геномной основы для расширения.
Материалы и методы
Растительный материал
C. makinoi Мацум. et Nakai или № JP131333 Растение Ryuunougiku, или для краткости C. makinoi , было получено из генного банка NARO (Цукуба, Япония). Черенки выращивали в теплицах Wageningen University and Research (WUR-Unifarm) по стандартным процедурам.
Экстракция ДНК, подготовка библиотеки и секвенирование
Высокомолекулярная ДНК для секвенирования с длительным считыванием была выделена из свежих молодых листьев C. makinoi с использованием модифицированного протокола (Bernatzky and Tanksley 1986). Библиотеки готовили с использованием наборов для секвенирования одномерного лигирования SQK-LSK108 и SQK-LSK109 (Oxford Nanopore Technologies, Оксфорд, Великобритания) в соответствии с инструкциями. Образцы секвенировали на приборе Oxford Nanopore GridION с использованием 40 проточных ячеек и стандартного протокола.Адаптеры были удалены с помощью Porechop (Wick 2018), а чтения были отфильтрованы с помощью Filtlong (Wick 2019), который удалил 10% худших чтений из более коротких чтений.
makinoi с использованием модифицированного протокола (Bernatzky and Tanksley 1986). Библиотеки готовили с использованием наборов для секвенирования одномерного лигирования SQK-LSK108 и SQK-LSK109 (Oxford Nanopore Technologies, Оксфорд, Великобритания) в соответствии с инструкциями. Образцы секвенировали на приборе Oxford Nanopore GridION с использованием 40 проточных ячеек и стандартного протокола.Адаптеры были удалены с помощью Porechop (Wick 2018), а чтения были отфильтрованы с помощью Filtlong (Wick 2019), который удалил 10% худших чтений из более коротких чтений.
Один образец был также секвенирован с использованием четырех библиотек вставок разного размера (270, 350, 400 и 500 п.н.) и чтения парных концов по 150 п.н. на Illumina HiSeq 2500 (GenomeScan, Лейден, Нидерланды). Образцы обрабатывали с использованием набора для подготовки библиотеки ДНК NEBNext ® Ultra от Illumina. Характеристики генома оценивали с помощью Jellyfish v2.2.10 (Marçais and Kingsford 2011) количество k-меров и GenomeScope (Vurture et al. 2017).
2017).
Высокомолекулярная ДНК C. makinoi также была секвенирована с помощью GenomeScan в восьми клетках SMRT с использованием секвенатора PacBio «Sequel SMRT Cell 1M v2». Подготовка образцов проводилась на основе протокола «PacBio SMRTbell Express Kit v1». Окончательная библиотека была выбрана с использованием протокола Blue Pippin для фрагментов размером более 15 kb. Праймер и полимеразу присоединяли с использованием набора «Sequel Binding and Internal Ctrl Kit2».1», и очистку проводили с использованием протокола PacBio «Процедура и контрольный список — очистка шариков AMPure ® PB комплексов SMRTbell ® , связанных с полимеразой». Секвенирование проводили в течение 10 часов на семи клетках и в течение 20 часов на остальных клетках с рекомендуемым количеством «Комплекса внутреннего контроля ДНК 2.1». Необработанные данные были оценены с помощью сервера SMRT Link Analysis v5.1.0.26367 GenomeScan.
Четыре ткани (листья, стебли, цветочные почки и цветки), использованные в исследовании, были получены из C. makinoi , выращенный в теплице в условиях длинного дня, 20-часовой световой день/4-часовой цикл темноты, или в условиях короткого дня, 11-часовой световой/13-часовой темновой цикл, в Dekker Chrysanten (Хенсбрук, Нидерланды) . Все собранные растительные ткани были немедленно заморожены в жидкости N 2 и хранились при -70°C до тех пор, пока РНК не была извлечена и выделена с использованием мини-набора RNeasy (Qiagen, Hilden, Германия) и библиотеки, подготовленной с использованием набора для секвенирования ПЦР-кДНК ( SQK-PCS109; Oxford Nanopore Technologies) в соответствии с инструкциями производителя.Образцы секвенировали отдельно на Oxford Nanopore GridION с использованием в общей сложности девяти проточных ячеек в соответствии со стандартным протоколом. Контроль качества осуществлялся с использованием NanoComp v1.9.2 (De Coster et al. 2018) и валидатора fastq из fastq_utils v0.21.0 (Fonseca and Manning) с удалением повторяющихся идентификаторов чтения.
makinoi , выращенный в теплице в условиях длинного дня, 20-часовой световой день/4-часовой цикл темноты, или в условиях короткого дня, 11-часовой световой/13-часовой темновой цикл, в Dekker Chrysanten (Хенсбрук, Нидерланды) . Все собранные растительные ткани были немедленно заморожены в жидкости N 2 и хранились при -70°C до тех пор, пока РНК не была извлечена и выделена с использованием мини-набора RNeasy (Qiagen, Hilden, Германия) и библиотеки, подготовленной с использованием набора для секвенирования ПЦР-кДНК ( SQK-PCS109; Oxford Nanopore Technologies) в соответствии с инструкциями производителя.Образцы секвенировали отдельно на Oxford Nanopore GridION с использованием в общей сложности девяти проточных ячеек в соответствии со стандартным протоколом. Контроль качества осуществлялся с использованием NanoComp v1.9.2 (De Coster et al. 2018) и валидатора fastq из fastq_utils v0.21.0 (Fonseca and Manning) с удалением повторяющихся идентификаторов чтения.
Сборка генома и каркас
чтения Nanopore были вызваны с помощью Guppy v3. 2 (Oxford Nanopore Technologies) и отфильтрованы, чтобы оставить только чтения из папки «проход» ( Q ≥ 7), длина которых превышает 20 КБ, и папки «сбой» ( Q < 7) длиной более 50 кб.В этот набор данных также были добавлены чтения PacBio длиной более 30 КБ. Эта комбинация длинных ридов была собрана с использованием SMARTdenovo v1.0.0 (Liu et al. 2021) с параметром «генерировать консенсус», установленным на 1. Затем была использована очистка гаплотигов (Roach et al. 2018), чтобы сгладить области гетерозиготности в единая согласованная последовательность. Данные Illumina впоследствии использовались в сочетании с ntEdit v0.9 (Warren et al. 2019) в режиме 2 и с K = 50 для двух итераций для полировки контигов.Смежность была дополнительно улучшена с использованием методов лигирования близости Hi-C и Чикаго (ласточкин хвост Genomics, Скоттс-Вэлли, США). Окончательный каркас на уровне псевдомолекул был выполнен с использованием ALLMAPS v0.
2 (Oxford Nanopore Technologies) и отфильтрованы, чтобы оставить только чтения из папки «проход» ( Q ≥ 7), длина которых превышает 20 КБ, и папки «сбой» ( Q < 7) длиной более 50 кб.В этот набор данных также были добавлены чтения PacBio длиной более 30 КБ. Эта комбинация длинных ридов была собрана с использованием SMARTdenovo v1.0.0 (Liu et al. 2021) с параметром «генерировать консенсус», установленным на 1. Затем была использована очистка гаплотигов (Roach et al. 2018), чтобы сгладить области гетерозиготности в единая согласованная последовательность. Данные Illumina впоследствии использовались в сочетании с ntEdit v0.9 (Warren et al. 2019) в режиме 2 и с K = 50 для двух итераций для полировки контигов.Смежность была дополнительно улучшена с использованием методов лигирования близости Hi-C и Чикаго (ласточкин хвост Genomics, Скоттс-Вэлли, США). Окончательный каркас на уровне псевдомолекул был выполнен с использованием ALLMAPS v0. 9.14 (Tang et al. 2015) и интегрированной генетической карты гексаплоидной хризантемы (van Geest et al. 2017a; см. дополнительную таблицу S1 и дополнительный рисунок S1). Также были внесены некоторые исправления неправильной сборки вручную, проверенные с помощью необработанных данных длительного чтения (см. Дополнительный рисунок S2).Контиги, которые остались не размещенными среди девяти хромосом в окончательной сборке, были отфильтрованы для удаления загрязнений и необычно высокого покрытия. Окончательные хромосомы были названы и пронумерованы в соответствии с назначением групп сцепления в C. morifolium Ramat. крест найден у van Geest et al. (2017а). Покрытие считывания оценивали с помощью Qualimap bamqc v2.2.1 (Оконечников и др. 2016), а контиги с отсутствием или высоким покрытием (более чем в 4 раза выше среднего) удаляли.Впоследствии последовательности загрязняющих веществ были идентифицированы с помощью Centrifuge v1.0.4 (Kim et al.
9.14 (Tang et al. 2015) и интегрированной генетической карты гексаплоидной хризантемы (van Geest et al. 2017a; см. дополнительную таблицу S1 и дополнительный рисунок S1). Также были внесены некоторые исправления неправильной сборки вручную, проверенные с помощью необработанных данных длительного чтения (см. Дополнительный рисунок S2).Контиги, которые остались не размещенными среди девяти хромосом в окончательной сборке, были отфильтрованы для удаления загрязнений и необычно высокого покрытия. Окончательные хромосомы были названы и пронумерованы в соответствии с назначением групп сцепления в C. morifolium Ramat. крест найден у van Geest et al. (2017а). Покрытие считывания оценивали с помощью Qualimap bamqc v2.2.1 (Оконечников и др. 2016), а контиги с отсутствием или высоким покрытием (более чем в 4 раза выше среднего) удаляли.Впоследствии последовательности загрязняющих веществ были идентифицированы с помощью Centrifuge v1.0.4 (Kim et al. 2016) с использованием вирусных и бактериальных библиотек NCBI (доступ в ноябре 2019 г.) и удалены. Остальные чтения были помещены в нулевую хромосому с N-промежутками в 200 п.н. между каждым контигом.
2016) с использованием вирусных и бактериальных библиотек NCBI (доступ в ноябре 2019 г.) и удалены. Остальные чтения были помещены в нулевую хромосому с N-промежутками в 200 п.н. между каждым контигом.
Органеллы были собраны путем извлечения прочтений Nanopore и Illumina, которые соответствовали доступным C. seticuspe (син. C. Boreale ), хлоропласту (Won et al. 2018b) и митохондриям (Won et al.. 2018c) ссылки с использованием Minimap2 v2.17 (Li 2018) и BWA-MEM v0.7.17-r1198-dirty (Li 2013) соответственно. Затем для каждой органеллы выполняли гибридную сборку с использованием Unicycler v0.4.8 (Wick et al. 2017). Это привело к образованию единого круглого каркаса для хлоропластов и множества круглых каркасов для митохондрий. На основании визуального осмотра каждого из каркасов митохондрий по сравнению с известными сборками митохондрий хризантем было обнаружено, что каркас 1 представляет всю последовательность и был выбран как полная круговая сборка митохондриального генома.
Геномный анализ и оценка качества
QUAST v5.0.2 (Gurevich et al. 2013) использовали для определения базовой статистики окончательной сборки генома, такой как общая длина, N50 и количество контигов/каркасов. BUSCO v4.0.5 (Simão et al. 2015) и соответствующий набор универсальных однокопийных ортологов Embryophyta odb10 также использовали для оценки полноты генома.
Повтор и расшифровка аннотации
Перед аннотированием сборки мы мягко замаскировали повторяющиеся последовательности с помощью RepeatModeler v2.0,1 (Флинн и др. 2020).
Предсказание генов было выполнено с помощью конвейера Funannotate v1.7.4 (Palmer 2017). Во-первых, конвейер Funannotate был обучен с использованием длинных чтений кДНК, базы данных UniProtKB v2020_04 (Bateman 2019) и базы данных белков BUSCO eukaryote odb9 (Simão et al. 2015), чтобы создать входной набор данных для конвейера прогнозирования Funannotate. Затем был запущен конвейер прогнозирования со стандартными настройками и алгоритмами GeneMark-ET, Augustus, GlimmerHMM и Snap.После этого была выполнена фильтрация предсказаний гена ab initio с использованием EVidenceModeler (EVM; Haas et al. 2008).
Затем был запущен конвейер прогнозирования со стандартными настройками и алгоритмами GeneMark-ET, Augustus, GlimmerHMM и Snap.После этого была выполнена фильтрация предсказаний гена ab initio с использованием EVidenceModeler (EVM; Haas et al. 2008).
Для функциональной аннотации предсказанных моделей было проведено первоначальное сравнение с использованием blastp v2.6.0 (Camacho et al. 2009) с базой данных SWISS-PROT v4 (Bairoch and Apweiler 2000) с отсечкой e — значение 1.0E−3, размер слова 6, максимальное количество попаданий установлено равным 20, и включен фильтр низкой сложности. Чтобы идентифицировать домены в наборах прогнозируемых моделей, InterProScan v5.26 (Jones et al. 2014) использовался вместе с библиотеками panther v12.0. Наконец, результаты были обработаны автономной версией Blast2Go (Götz и др. 2008 г.) с настройками по умолчанию.
Результаты и обсуждение
Качество необработанной последовательности
В результате секвенирования Nanopore было получено 443,25 ГБ данных с числом чтения N50, равным 22,6 КБ. После базового вызова, удаления адаптеров и фильтрации чтений длиной более 20 КБ из папки «проход», которая имела оценку Q > 7, и чтений длиной более 50 КБ из папки «не пройдено», набор данных имел покрытие примерно 53× (при условии, что размер гаплоидного генома равен 3.1 ГБ) и состоял из 3 924 770 чтений. Illumina HiSeq предоставила 113,2, 142,0, 133,7 и 120,0 Гб необработанных данных для библиотек размера вставки 270, 350, 400 и 500 п.н. соответственно. От 90,5% до 94,6% прочтений для каждого размера вставки имели оценку качества « q », большую или равную 30. В результате секвенирования PacBio было получено 70 ГБ данных со средней длиной подчтения 15,5 кБ и N50, равным 24,1. кб. Это означало охват примерно 30,6× (при условии, что размер гаплоидного генома составляет 3,1 ГБ).
После базового вызова, удаления адаптеров и фильтрации чтений длиной более 20 КБ из папки «проход», которая имела оценку Q > 7, и чтений длиной более 50 КБ из папки «не пройдено», набор данных имел покрытие примерно 53× (при условии, что размер гаплоидного генома равен 3.1 ГБ) и состоял из 3 924 770 чтений. Illumina HiSeq предоставила 113,2, 142,0, 133,7 и 120,0 Гб необработанных данных для библиотек размера вставки 270, 350, 400 и 500 п.н. соответственно. От 90,5% до 94,6% прочтений для каждого размера вставки имели оценку качества « q », большую или равную 30. В результате секвенирования PacBio было получено 70 ГБ данных со средней длиной подчтения 15,5 кБ и N50, равным 24,1. кб. Это означало охват примерно 30,6× (при условии, что размер гаплоидного генома составляет 3,1 ГБ).
В результате секвенирования кДНК с нанопорами были получены наборы данных с 4.8–7,9 млн операций чтения, средний размер N50 составляет 1,2–1,4 КБ, а общий объем составляет от 5,0 до 7,9 ГБ (таблица 1).
Детали секвенирования кДНК из различных органов растений в C. makinoi
| Источник . | Средняя длина считывания (б) . | Среднее качество чтения . | Количество чтений . | Длина считывания N50 (б) . | Всего баз .+ | |
|---|---|---|---|---|---|---|
| лист (короткий г) | 977,0 | 9,0 | 7587930 | 1 189 | 7413732394 | |
| лист (длинный д) | 973,0 | 9,4 | 6780899 | 1209 | 6597664366 | |
| Calyx | ||||||
| Calyx | 10404 | 10.0 | 4 8333930 | 4 833397 | 1 247 9 | 5 027 548 849 |
| Цветочный дисковый флонты | 1 012.1 | 9,5 | 7072131 | 1331 | 71575 | |
| Цветочные почки | 993,6 | 9,1 | 70 | тысяча двести пятьдесят шесть | 7867489497 | |
| Цветок луч цветочки | 1,002. 8 8 | 9,6 | 7000372 | 1,250 | 7 020311 566 | |
| Meristem | 1 048,0 | 1 04 | 5 075,164 | 1 263 | 5 318 809662 | |
| Стебель (короткие д) | 997.1 | 9.5 7 3 | тысячу двести тридцать две 7 | 3286 | ||
| Корень | 1060 | 8,4 5272384 | +1389 55 | 404 |
| Источник . | Средняя длина считывания (б) . | Среднее качество чтения . | Количество чтений . | Длина считывания N50 (б) . | Всего баз . + | |
|---|---|---|---|---|---|---|
| лист (короткий г) | 977,0 | 9,0 | 7587930 | 1 189 | 7413732394 | |
| лист (длинный д) | 973,0 | 9,4 | 6780899 | 1209 | 6597664366 | |
| Calyx | ||||||
| Calyx | 10404 | 10. 0 0 | 4 8333930 | 4 833397 | 1 247 9 | 5 027 548 849 |
| Цветочный дисковый флонты | 1 012.1 | 9,5 | 7072131 | 1331 | 71575 | |
| Цветочные почки | 993,6 | 9,1 | 70 | тысяча двести пятьдесят шесть | 7867489497 | |
| Цветок луч цветочки | 1,002.8 | 9,6 | 7000372 | 1,250 | 7 020311 566 | |
| Meristem | 1 048,0 | 1 04 | 5 075,164 | 1 263 | 5 318 809662 | |
| Стебель (короткие д) | 997.1 | 9,5 | 7 3 | 1232 | 7 | 3286 |
| Корень | +1060 | 8,4 | 5272384 | тысяча триста восемьдесят-девять | 55 | 404 |
секвенирования детали кДНК из различных органов растений в C. makinoi
makinoi
| Источник . | Средняя длина считывания (б) . | Среднее качество чтения . | Количество чтений . | Длина считывания N50 (б) . | Всего баз . + | |||
|---|---|---|---|---|---|---|---|---|
| лист (короткий г) | 977,0 | 9,0 | 7587930 | 1189 | 7413732394 | |||
| лист (длинный д) | 973,0 | 9,4 | 6780899 | 1209 | 6597664366 | |||
| Чашечка | 1040.2 | 10,0 | 4833397 | 1247 | 5027548849 | |||
| Цветочный диск цветочки | 1,012.1 | 9,5 | 7072131 | 1331 | 71575 | |||
| Цветочные почки | 993,6 | 9,1 | 70 | 1,256 | 1,256 | 7 867 489497 | ||
| цветок лучей флонты | 1,002. 8 8 | 9.6 | 7 000 372 | 7 000 372 | 1250 | 7 020 311 566660930 | ||
| Meristem | 1 048.0 | 10,2 | 5075164 | 1 263 | 5318808662 | |||
| Стебель (короткий г) | 997,1 | 9,5 | 7 3 | тысячу двести тридцать-две | 7 | 3286 | ||
| Корень | +1060 | 8,4 | Источник . | Средняя длина считывания (б) . | Среднее качество чтения . | Количество чтений . | Длина считывания N50 (б) . | Всего баз . + |
| лист (короткий г) | 977,0 | 9,0 | 7587930 | 1189 | 7413732394 | |||
| лист (длинный д) | 973,0 | 9,4 | 6780899 | 1209 | 6597664366 | |||
| Чашечка | 1040. 2 2 | 10,0 | 4833397 | 1247 | 5027548849 | |||
| Цветочный диск цветочки | 1,012.1 | 9,5 | 7072131 | 1331 | 71575 | |||
| Цветочные почки | 993,6 | 9,1 | 70 | 1,256 | 1,256 | 7 867 489497 | ||
| цветок лучей флонты | 1,002.8 | 9.6 | 7 000 372 | 7 000 372 | 1250 | 7 020 311 566660930 | ||
| Meristem | 1 048.0 | 10,2 | 5075164 | 1263 | 5318808662 | |||
| Стебель (короткий г) | 997,1 | 9,5 | 7 3 | тысячу двести тридцать два | 7 | 3286 | ||
| Корень | +1060 | 8,4 | 5 272 384 | 1 389 | 5 591 241 404 |
Размер и характеристики генома
k-меров ( K = 31) были извлечены из парных концевых прочтений HiSeq Illumina, подсчитанных с использованием Jellyfish v2. 2.10 (Marçais and Kingsford 2011) и проанализированы с помощью GenomeScope (Vurture et al. 2017) для оценки гаплоидной длины генома, гетерозиготности и содержания повторов. Анализ сошелся и оценил размер гаплоидного генома в 1,72 Gb, гетерозиготность 1,51% (это значение колеблется от ~0% до 2% (Vurture et al. 2017)) и пометило 53,6% генома как уникальное (рис. 1). Это указывает на повторяемость генома и высокую гетерозиготность. Размер гаплоидного генома диплоидов хризантем оценивается между 2.90 ± 0,03 Gb для C. seticuspe (Hirakawa et al. 2019) и 3,24 Gb для C. nankingense (Song et al. 2018) с использованием проточной цитометрии. Предыдущая оценка размера генома C. makinoi (Nakano et al. 2019) предполагала, что геном был примерно на 10% больше, чем C. seticuspe , или примерно 3,19 ГБ. База данных размера генома Asteraceae оценивает средний 1C в 3,82 Гб для хризантемы с использованием проточной цитометрии, хотя это, вероятно, завышенная оценка, поскольку медиана составляет 3.
2.10 (Marçais and Kingsford 2011) и проанализированы с помощью GenomeScope (Vurture et al. 2017) для оценки гаплоидной длины генома, гетерозиготности и содержания повторов. Анализ сошелся и оценил размер гаплоидного генома в 1,72 Gb, гетерозиготность 1,51% (это значение колеблется от ~0% до 2% (Vurture et al. 2017)) и пометило 53,6% генома как уникальное (рис. 1). Это указывает на повторяемость генома и высокую гетерозиготность. Размер гаплоидного генома диплоидов хризантем оценивается между 2.90 ± 0,03 Gb для C. seticuspe (Hirakawa et al. 2019) и 3,24 Gb для C. nankingense (Song et al. 2018) с использованием проточной цитометрии. Предыдущая оценка размера генома C. makinoi (Nakano et al. 2019) предполагала, что геном был примерно на 10% больше, чем C. seticuspe , или примерно 3,19 ГБ. База данных размера генома Asteraceae оценивает средний 1C в 3,82 Гб для хризантемы с использованием проточной цитометрии, хотя это, вероятно, завышенная оценка, поскольку медиана составляет 3. 1 Gb (Garnatje и др. 2011). Известно, что методы оценки генома на основе последовательностей недооценивают размер генома (Pflug et al. 2020), при этом GenomeScope особенно чувствителен к параметру отсечки количества k-mer (Vurture et al. 2017). Этот параметр предназначен для того, чтобы отличать повторяющиеся последовательности от последовательностей органелл, так что повторяющиеся k-меры используются для расчета размера генома, в то время как k-меры органелл отбрасываются, но это становится невозможным, если повторяющиеся k-меры последовательностей столь же многочисленны, как и k-меры органелл. k-меры органелл (Vurture et al. 2017). При высоком уровне гетерозиготности, показанном GenomeScope и подтвержденном более поздними анализами, было бы трудно отличить эти k-меры друг от друга, что привело к тому, что многие k-меры повторяющихся областей также были отброшены, что привело к существенно недооценке размера генома. Мы ожидаем, что истинный размер генома будет ближе к предыдущим прогнозам цитометрии, составляющим 3,19 Гб (Nakano et al.
1 Gb (Garnatje и др. 2011). Известно, что методы оценки генома на основе последовательностей недооценивают размер генома (Pflug et al. 2020), при этом GenomeScope особенно чувствителен к параметру отсечки количества k-mer (Vurture et al. 2017). Этот параметр предназначен для того, чтобы отличать повторяющиеся последовательности от последовательностей органелл, так что повторяющиеся k-меры используются для расчета размера генома, в то время как k-меры органелл отбрасываются, но это становится невозможным, если повторяющиеся k-меры последовательностей столь же многочисленны, как и k-меры органелл. k-меры органелл (Vurture et al. 2017). При высоком уровне гетерозиготности, показанном GenomeScope и подтвержденном более поздними анализами, было бы трудно отличить эти k-меры друг от друга, что привело к тому, что многие k-меры повторяющихся областей также были отброшены, что привело к существенно недооценке размера генома. Мы ожидаем, что истинный размер генома будет ближе к предыдущим прогнозам цитометрии, составляющим 3,19 Гб (Nakano et al. 2019).
2019).
Рисунок 1
Распределение k-mer ( K = 31) на основе ридов Illumina, смоделированных и визуализированных с помощью геномоскопа.
Рисунок 1
Распределение k-mer ( K = 31) на основе показаний illumina, смоделированных и визуализированных с помощью геномоскопа.
Сборка и качество генома
После первоначальной сборки с помощью SMARTdenovo (Ruan и др. , 2017 г.) у нас было 39 105 контигов размером 4,1 ГБ с N50 размером 139,2 КБ. Purge Haplotigs (Roach et al. 2018) создал плоскую сборку из 15 236 контигов, охватывающую 3,1 Гб, с N50 размером 255,8 КБ.После двух раундов полировки с помощью ntEdit v0.9 (Warren et al. 2019) с использованием данных Illumina размер сборки составил 3,1 Гб и состоял из 15 226 контигов, а N50 — 258,2 КБ.
Для создания каркаса контигов были созданы карты с использованием методов Hi-C и чикагского лигирования. Этот метод сгенерировал 4254 скаффолда общей длиной 3,1 Гб с N50 168,9 Мб. Эта сборка была дополнительно преобразована в псевдохромосомы с использованием ALLMAPS v0.9.14 (Tang et al. 2015) с использованием генетической карты гексаплоида C. moriflorium Ramat. (ван Геест и др. 2017a). В результате была получена окончательная сборка длиной 3,1 Гб, состоящая из девяти псевдохромосом с N50 и L50 размером 330,0 Мб и пяти каркасов соответственно (таблица 2).
Эта сборка была дополнительно преобразована в псевдохромосомы с использованием ALLMAPS v0.9.14 (Tang et al. 2015) с использованием генетической карты гексаплоида C. moriflorium Ramat. (ван Геест и др. 2017a). В результате была получена окончательная сборка длиной 3,1 Гб, состоящая из девяти псевдохромосом с N50 и L50 размером 330,0 Мб и пяти каркасов соответственно (таблица 2).
C. makinoi de novo , оцененные с использованием QUAST
| Сборка . | С.makinoi V1.0 (9 chrs + chr0) . 90 914 | |
|---|---|---|
| # Ns на 100 т.п.н. | 89,51 | |
| # контигов / каркасы | 10 | |
| Общая длина | 3113668257 | |
| N50 | 330012911 | |
| N75 | 317988395 | |
| L50 | ||
| 5 | 5 | |
| L75 | 7 | 7 |
| Крупнейшие наряды / леса | 376 468 909 | 0 |
| GC Content (%) | 36. 01 01 |
| Сборка . | C. makinoi V1.0 (9 хр + хр0) . 90 914 | |
|---|---|---|
| # Ns на 100 т.п.н. | 89,51 | |
| # контигов / каркасы | 10 | |
| Общая длина | 3113668257 | |
| N50 | 330012911 | |
| N75 | 317988395 | |
| L50 | ||
| 5 | 5 | |
| L75 | 7 | 7 |
| Крупнейшие наряды / леса | 376 468 909 | 0 |
| GC Content (%) | 36.01 |
Показатели сборки генома C. makinoi de novo , рассчитанные с использованием QUAST
| Сборка . | C. makinoi V1.0 (9 хр + хр0) . 90 914 | |
|---|---|---|
| # Ns на 100 т.п.н. | 89,51 | |
| # контигов / каркасы | 10 | |
| Общая длина | 3113668257 | |
| N50 | 330012911 | |
| N75 | 317988395 | |
| L50 | ||
| 5 | 5 | |
| L75 | 7 | 7 |
| Крупнейшие наряды / леса | 376 468 909 | 0 |
| GC Content (%) | 36. 01 01 |
| Сборка . | C. makinoi V1.0 (9 хр + хр0) . 90 914 | |
|---|---|---|
| # Ns на 100 т.п.н. | 89,51 | |
| # контигов / каркасы | 10 | |
| Общая длина | 3113668257 | |
| N50 | 330012911 | |
| N75 | 317988395 | |
| L50 | ||
| 5 | 5 | |
| L75 | 7 | 7 |
| Крупнейшие наряды / леса | 376 468 909 | 0 |
| GC Content (%) | 36.01 |
Неразмещенные контиги были отобраны перед помещением в хромосому 0 с использованием механизма классификации Centrifuge v1.0.4 (Kim et al. 2016). Из 4206 неразмещенных контигов 824 были помечены как происходящие из неэукариотического источника и удалены. Прочтения Illumina также были выровнены по всем контигам с помощью Minimap2 v2. 17 (Li 2018), а затем их покрытие было оценено с помощью Qualimap v2.2.1 (Okonechnikov et al. 2016). Контиги с покрытием выше, чем одно стандартное отклонение от среднего, были удалены.В результате был получен окончательный набор из 3337 контигов, покрывающих в общей сложности 198,3 Mb, которые были помещены в хромосому 0.
17 (Li 2018), а затем их покрытие было оценено с помощью Qualimap v2.2.1 (Okonechnikov et al. 2016). Контиги с покрытием выше, чем одно стандартное отклонение от среднего, были удалены.В результате был получен окончательный набор из 3337 контигов, покрывающих в общей сложности 198,3 Mb, которые были помещены в хромосому 0.
балла BUSCO, которые представляют собой набор универсальных однокопийных ортологов, также использовались для оценки полноты сборок. (см. Таблицу 3). При использовании набора Embryophyta odb10 с BUSCO v4.0.5 (Simão et al. 2015) окончательная сборка получила полный балл BUSCO 92,1%, что указывает на высокое общее качество. Полную разбивку баллов BUSCO можно увидеть в таблице 3.
Таблица 3Результаты конвейера анализа Busco для оценки полноты набора генов
| Термин BUSCO . | V1.0 . | ||
|---|---|---|---|
| Complete (%) | 92.1 | 92. 1 1 | |
| Полная и одиночная копия (%) | 83.8 | ||
| Комплектация и дублированные (%) | 8.3 | ||
| Фрагментированные (%) | 1.8 | ||
| Отсутствует (%) | 6.1 | ||
| Всего | 1375 | 912 BCO термин 3 | 909 909 .V1.0 . |
| Complete (%) | 92.1 | 92.1 | |
| Полная и одиночная копия (%) | 83.8 | ||
| Комплектация и дублированные (%) | 8.3 | ||
| Фрагментированные (%) | 1.8 | ||
| отсутствуют (%) | 6.1 | ||
| Всего | 1375 | 1375 | 0 |
Выход из анализа Busco Consure для оценки Gene Complete Complete
12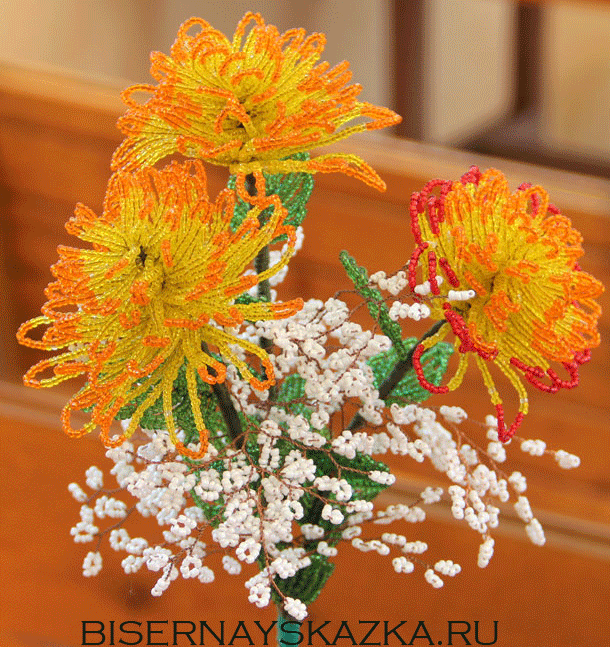 0
.
0
.| Terms . | V1.0 . | |
|---|---|---|
| Полное (%) | 92.1 | 92.1 |
| Полная и одиночная копия (%) | 83.8 | |
| Полное и дублированное (%) | 8.3 | |
| Фрагментированные (%) | 1.8 | |
| Отсутствуют (%) | 6.1 | |
| Всего | 1375 | 0 |
Для сравнения, исключительно вспомогательное собрание на основе C . seticuspe имел общую длину 2,722 Гб, 354 212 контигов, N50 44,7 тб и оценку BUSCO 88,8% (Hirakawa et al. 2019). Сборка C. nankingense имела общую длину 2 м.527 ГБ, с 24 051 контигом, N50 – 130,7 КБ и оценкой BUSCO – 92,7% (Song et al. 2018) . Таким образом, мы смогли создать значительно более непрерывную сборку без ущерба для полноты.
seticuspe имел общую длину 2,722 Гб, 354 212 контигов, N50 44,7 тб и оценку BUSCO 88,8% (Hirakawa et al. 2019). Сборка C. nankingense имела общую длину 2 м.527 ГБ, с 24 051 контигом, N50 – 130,7 КБ и оценкой BUSCO – 92,7% (Song et al. 2018) . Таким образом, мы смогли создать значительно более непрерывную сборку без ущерба для полноты.
Повторяющиеся области
Используя RepeatModeler (Flynn et al. 2020), 80,04% генома были отмечены как повторяющиеся. Большие геномы накапливают повторы (Kelly and Leitch, 2011), и анализ k-mer уже показал, что мы имеем дело с в значительной степени повторяющимся геномом.Из 6799 идентифицированных семейств повторов у C. makinoi 76,6% были идентифицированы как длинные концевые повторы (LTR). Из LTR 27,1% могут быть идентифицированы как Copia и 7,4% как Gypsy . Аналогичный анализ C. nankingense пометил 69,6% их сборки как повторяющиеся и обнаружил, что LTR составляют 67,7% идентифицированных тандемных повторов, при этом 36,5% составляют Copia и 30,9% составляют Gypsy (Song et al. 2018). Меньший уровень повторяемости и идентифицированные LTR в 90–201 C.nankingense может быть связано с разницей в смежности: C. nankingense состоит из более чем 24000 контигов (Song et al. 2018) с нашими 9 псевдохромосомами и 3337 неразмещенными контигами, поскольку было показано, что более полные сборки генома будет идентифицировать больше LTR (Ou и др. 2018). Анализ различных сложноцветных показал колебания относительной численности Copia по сравнению с Gypsy , при этом подсолнечник ( Helianthus annuus ) усиливает Gyspy по сравнению с Copia (Cavallini et al.. 2010; Бути и др. 2011; Натали и др. 2013; Джордани и др. 2014; Бадуин и др. 2017), в то время как конский сорняк ( Conyza canadensis ) и артишок шаровидный ( Cynara Cardunculus var. scolymus) показали обратное (Peng et al. 2014; Scaglione et al.
2018). Меньший уровень повторяемости и идентифицированные LTR в 90–201 C.nankingense может быть связано с разницей в смежности: C. nankingense состоит из более чем 24000 контигов (Song et al. 2018) с нашими 9 псевдохромосомами и 3337 неразмещенными контигами, поскольку было показано, что более полные сборки генома будет идентифицировать больше LTR (Ou и др. 2018). Анализ различных сложноцветных показал колебания относительной численности Copia по сравнению с Gypsy , при этом подсолнечник ( Helianthus annuus ) усиливает Gyspy по сравнению с Copia (Cavallini et al.. 2010; Бути и др. 2011; Натали и др. 2013; Джордани и др. 2014; Бадуин и др. 2017), в то время как конский сорняк ( Conyza canadensis ) и артишок шаровидный ( Cynara Cardunculus var. scolymus) показали обратное (Peng et al. 2014; Scaglione et al. 2016). Более ранние исследования с C. nankingense и C. seticuspe (син. C. Boreale ) показали, что в хризантемах распространенность Copia и Gypsy была сходной, при этом Copia была немного более многочисленной и подвергалась воздействию . усиление чуть раньше (Song et al. 2018; Вон и др. 2018a), но наши результаты показывают, что, по крайней мере, у C. makinoi существует более существенная разница в численности, как это наблюдается у конского сорняка и артишока шаровидного. Для лучшего понимания следует провести систематический анализ различных видов хризантем на различных уровнях плоидности, поскольку эти повторяющиеся типы являются известной движущей силой эволюции генома растений (Todorovska 2007).
2016). Более ранние исследования с C. nankingense и C. seticuspe (син. C. Boreale ) показали, что в хризантемах распространенность Copia и Gypsy была сходной, при этом Copia была немного более многочисленной и подвергалась воздействию . усиление чуть раньше (Song et al. 2018; Вон и др. 2018a), но наши результаты показывают, что, по крайней мере, у C. makinoi существует более существенная разница в численности, как это наблюдается у конского сорняка и артишока шаровидного. Для лучшего понимания следует провести систематический анализ различных видов хризантем на различных уровнях плоидности, поскольку эти повторяющиеся типы являются известной движущей силой эволюции генома растений (Todorovska 2007).
Аннотация стенограммы
Каждый алгоритм в конвейере Funannotate (Palmer 2017) создал набор ab initio моделей генов (см. Дополнительную таблицу S2). Доказательства для каждой модели гена были взвешены с использованием подхода EVM и идентифицированы 95 064 90 201 ab initio 90 202 предсказанных моделей генов. Это выше, чем в среднем по растениям (36 795) (Рамирес-Санчес и др. , 2016 г.), но может быть объяснено наличием псевдогенов (Сяо и др. , 2016 г.). Другие сложноцветные, включая Artemisia annua (63 226 моделей генов; Shen et al. 2018), подсолнечник (52 232 модели генов; Badouin et al. 2017), Mikania micrantha (46 1. 351 модель гена Liu et al2; 2020 г.), C. seticuspe (71 057 генных моделей; Hirakawa et al. 2019 г.) и C. nankingense (56 870 генных моделей; Song et al. 2018 г.) моделей генов. Для дальнейшего исследования также был проведен анализ структуры и длины аннотированных генов. Гены имели среднюю длину кодирующей последовательности 876 п.н. и максимальную 12 735 п.н. Это меньше, чем средняя длина гена растения, составляющая 1308 п.
Доказательства для каждой модели гена были взвешены с использованием подхода EVM и идентифицированы 95 064 90 201 ab initio 90 202 предсказанных моделей генов. Это выше, чем в среднем по растениям (36 795) (Рамирес-Санчес и др. , 2016 г.), но может быть объяснено наличием псевдогенов (Сяо и др. , 2016 г.). Другие сложноцветные, включая Artemisia annua (63 226 моделей генов; Shen et al. 2018), подсолнечник (52 232 модели генов; Badouin et al. 2017), Mikania micrantha (46 1. 351 модель гена Liu et al2; 2020 г.), C. seticuspe (71 057 генных моделей; Hirakawa et al. 2019 г.) и C. nankingense (56 870 генных моделей; Song et al. 2018 г.) моделей генов. Для дальнейшего исследования также был проведен анализ структуры и длины аннотированных генов. Гены имели среднюю длину кодирующей последовательности 876 п.н. и максимальную 12 735 п.н. Это меньше, чем средняя длина гена растения, составляющая 1308 п. н., но находится в пределах первого квартиля средней длины гена растения (Ramirez-Sánchez et al. 2016). В соответствии с выводом о том, что у растений, как правило, меньше экзонов на белок, чем у других организмов (Рамирес-Санчес и др. 2016), было обнаружено, что 15,9% генов в C. makinoi состоят из одного экзона. Было обнаружено, что средняя длина интрона в нашем наборе генов составляет 446 п.н. с диапазоном 11–19 668 п.н. и медианой 140 п.н. Это указывает на то, что большинство интронов относительно малы. Распределение похоже на то, что было обнаружено у кукурузы (у которой было среднее значение 516 п.н. и медиана 146 п.н.; Schnable et al. 2009).
н., но находится в пределах первого квартиля средней длины гена растения (Ramirez-Sánchez et al. 2016). В соответствии с выводом о том, что у растений, как правило, меньше экзонов на белок, чем у других организмов (Рамирес-Санчес и др. 2016), было обнаружено, что 15,9% генов в C. makinoi состоят из одного экзона. Было обнаружено, что средняя длина интрона в нашем наборе генов составляет 446 п.н. с диапазоном 11–19 668 п.н. и медианой 140 п.н. Это указывает на то, что большинство интронов относительно малы. Распределение похоже на то, что было обнаружено у кукурузы (у которой было среднее значение 516 п.н. и медиана 146 п.н.; Schnable et al. 2009).
Другим объяснением того, что предсказанные гены являются более многочисленными, является то, что среднее значение у растений может быть связано с дупликацией древнего генома. Ранее сообщалось, что до разделения между астеридами I и II, примерно 122–164 млн лет назад, у двудольных произошло древнее трипликация всего генома (WGT-γ) и еще одно трипликация всего генома (WGT-1). 53–62 млн лет назад (Badouin et al. 2017; Won et al. 2017; Liu et al. 2020). Кроме того, анализ частот синонимических замен паралогичных и ортологических генов транскриптомных сборок гексаплоида C. morifolium Ramat. и дикий корейский диплоид C. Boreale выявил событие дупликации или трипликации всего генома, специфичное для хризантемы (Won et al. 2017). Сборка и аннотация более качественных геномов хризантем помогут прояснить эволюцию рода и его вклад в обилие генов.
53–62 млн лет назад (Badouin et al. 2017; Won et al. 2017; Liu et al. 2020). Кроме того, анализ частот синонимических замен паралогичных и ортологических генов транскриптомных сборок гексаплоида C. morifolium Ramat. и дикий корейский диплоид C. Boreale выявил событие дупликации или трипликации всего генома, специфичное для хризантемы (Won et al. 2017). Сборка и аннотация более качественных геномов хризантем помогут прояснить эволюцию рода и его вклад в обилие генов.
Обычно мобильные элементы накапливаются в центромерной и перицентромерной областях, поскольку они устанавливают, поддерживают и стабилизируют центромеры эукариот (Klein and O’Neill 2018). Таким образом, можно оценить центромерную область хромосомы на основе низкой плотности генов (рис. 2; красное кольцо) и высокой плотности повторяющихся последовательностей (рис. 2; оранжевое кольцо), но этот паттерн не виден у C. makinoi , поскольку оба гены (красное кольцо) и повторяющиеся последовательности (оранжевое кольцо) равномерно распределены. Фактически, вместо кластеризации по регионам плотность повторяющихся последовательностей в C. makinoi имеет положительное значение корреляции Пирсона 0,60 с плотностью генов. Возможное объяснение этой корреляции заключается в том, что у хризантем, как и у других сложноцветных, есть LTR, обусловливающие большое разнообразие (Wang et al. 2014). Каждое семейство LTR имеет свои особенности распределения в геномах растений (Chen 2007; Zhang et al. 2014), и LTR составляют 76,6% идентифицированных семейств повторов у C.макиной . Огромный объем LTR, которые распределяются в богатых генами областях, может подавлять сигнал повторяющихся последовательностей с центромерным/перицентромерным предпочтением. Это также подтверждается предыдущей работой по повторяющимся элементам в C. seticuspe (син. C. Boreale ), которая с использованием оптических методов обнаружила сильное обогащение LTR и что большинство идентифицированных повторяющихся последовательностей не показали предпочтение центромерных или перицентромерных областей (Won et al.
Фактически, вместо кластеризации по регионам плотность повторяющихся последовательностей в C. makinoi имеет положительное значение корреляции Пирсона 0,60 с плотностью генов. Возможное объяснение этой корреляции заключается в том, что у хризантем, как и у других сложноцветных, есть LTR, обусловливающие большое разнообразие (Wang et al. 2014). Каждое семейство LTR имеет свои особенности распределения в геномах растений (Chen 2007; Zhang et al. 2014), и LTR составляют 76,6% идентифицированных семейств повторов у C.макиной . Огромный объем LTR, которые распределяются в богатых генами областях, может подавлять сигнал повторяющихся последовательностей с центромерным/перицентромерным предпочтением. Это также подтверждается предыдущей работой по повторяющимся элементам в C. seticuspe (син. C. Boreale ), которая с использованием оптических методов обнаружила сильное обогащение LTR и что большинство идентифицированных повторяющихся последовательностей не показали предпочтение центромерных или перицентромерных областей (Won et al. 2018а).
2018а).
Рисунок 2
График Circos, показывающий псевдомолекулы (внешнее кольцо), плотность генов (красное кольцо), содержание повторяющихся элементов (оранжевое кольцо) и содержание gc (желтое кольцо) с размером ячейки 500 кб.
Рисунок 2
График Circos, показывающий псевдомолекулы (внешнее кольцо), плотность генов (красное кольцо), содержание повторяющихся элементов (оранжевое кольцо) и содержание gc (желтое кольцо) с размером ячейки 500 kb.
Blast2Go (Götz и др. 2008) использовали для функциональной аннотации окончательного набора моделей генов. Из наших предсказанных моделей генов 11,0% были присвоены предполагаемые функциональные метки, а 2,9% — коды ферментов. Глядя на распределение на уровне GO, большинство моделей генов, которые были аннотированы как относящиеся к биологическому процессу (P) или молекулярной функции (F), не могли быть идентифицированы с высоким уровнем специфичности, за исключением клеточного компонента (C). аннотированные гены (см. Дополнительную фигуру S3). Это означает, что Blast2Go изо всех сил пытался уточнить функцию идентифицированных генов биологических процессов за пределами i.е. , «метаболический процесс соединения азота», но может быть гораздо более конкретным с аннотированными генами клеточного компонента.
аннотированные гены (см. Дополнительную фигуру S3). Это означает, что Blast2Go изо всех сил пытался уточнить функцию идентифицированных генов биологических процессов за пределами i.е. , «метаболический процесс соединения азота», но может быть гораздо более конкретным с аннотированными генами клеточного компонента.
Заключение
Собрав наиболее полный и непрерывный геном хризантем, доступный на сегодняшний день, мы сделали важный шаг вперед в нашем понимании геномики этой сложной и важной декоративной культуры. Этот справочник послужит руководством для дальнейших исследований селекционных признаков хризантем, их происхождения и стратегий создания родственных сортов с более высокой плоидностью.Этот геном может служить эталоном для помощи в упорядочении последовательностей других диплоидных хризантем, а также для уменьшения сложности сборки близкородственных полиплоидов, как это было сделано для нескольких других видов (Lukaszewski et al. 2014; Li ). и др. 2015 г.; Бертиоли и др. 2016 г.; Кириакиду и др. 2018 г.; Эджер и др. 2019 г.).
2014; Li ). и др. 2015 г.; Бертиоли и др. 2016 г.; Кириакиду и др. 2018 г.; Эджер и др. 2019 г.).
Доступность данных
Окончательная сборка и файлы аннотаций для C.makinoi Мацум. et Nakai (японское название: Ryuunougiku) № JP131333 доступны для загрузки на сайте www.chrysanthemumgenome.wur.nl/ вместе с браузером генома. Все необработанные данные, а также файлы сборки и аннотаций также можно найти в ENA под номером PRJEB44800. Образец растения доступен в Генбанке НАРО.
Дополнительный материал доступен по адресу G3 в Интернете.
Благодарности
Мы хотели бы поблагодарить Национальную сельскохозяйственную и продовольственную организацию (Япония) за то, что она предоставила нам образец растения Chrysanthemum makinoi JP131333 для этого проекта.
Финансирование
Мы хотели бы поблагодарить наших отраслевых партнеров, Deliflor Chrysanten, Dekker Chrysanten, Dümmen Orange и Royal Van Zanten, а также TKI T&U за их финансирование и поддержку в рамках проекта TKI KV1605-114.
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов.
Ссылки на литературу
Акерсон
К.
1967
.Оригинальный вид хризантемы
.Natl Chrysanth Soc Bull
.23
:105
–107
.Андерсон
№.
2007
. Хризантема. В кн.:Селекция цветов и генетика
.Спрингер
. п.389
–437
. Дой: 10.1007/978-1-4020-4428-1_14.Badouin
H
,Gouzy
J
,Grassa
CJ
,Murat
F
,Staton
SE
, et al.2017
.Геном подсолнечника дает представление о метаболизме масла, цветении и эволюции астерид
.Природа
.546
:148
–152
.Байрох
А
,Апвайлер
Р.
2000
.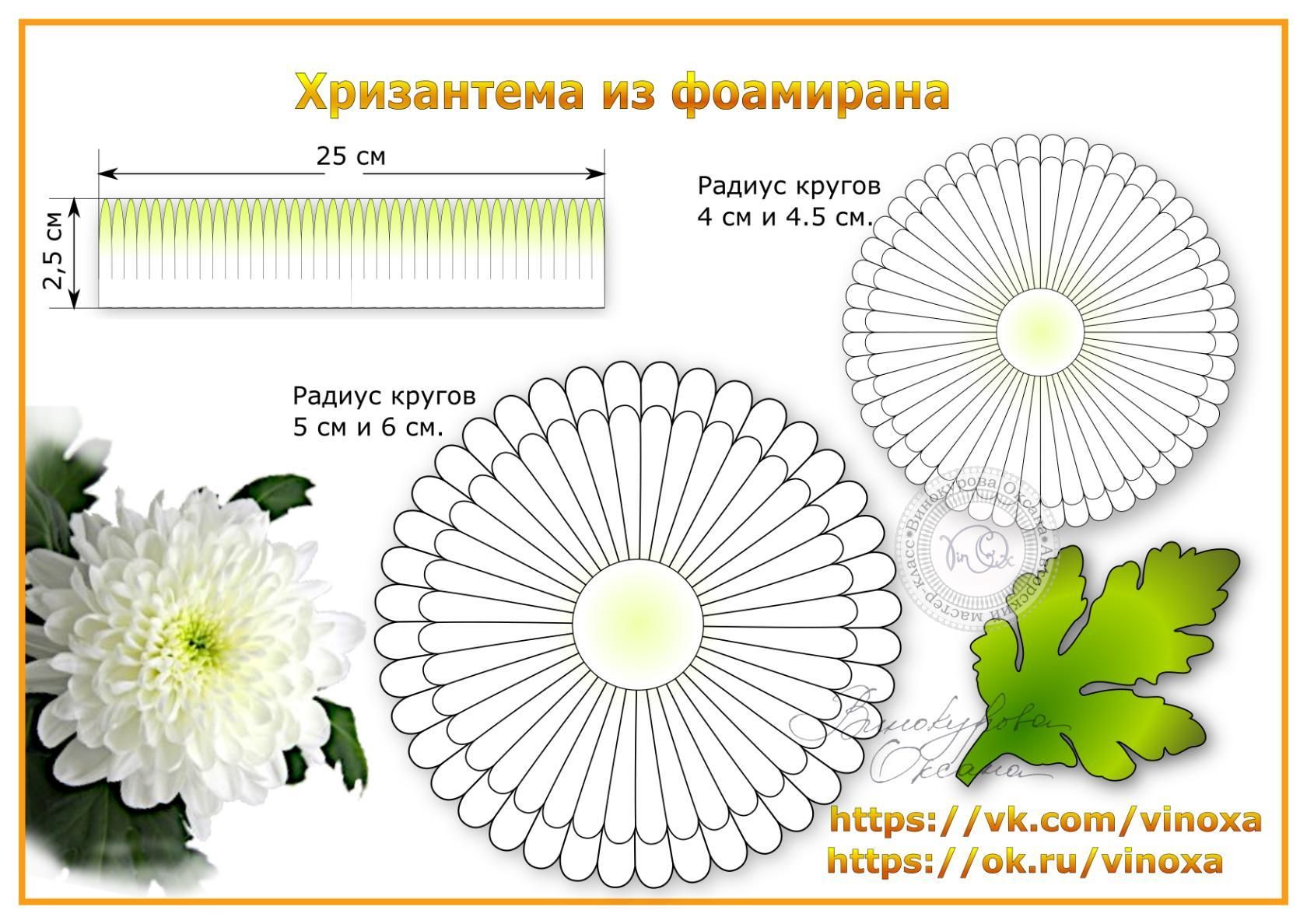
База данных белковых последовательностей SWISS-PROT и дополнение к ней TrEMBL в 2000 г.
.Рез. нуклеиновых кислот
.28
:45
–48
.Бейтман
А.
2019
.UniProt: всемирный центр знаний о белках
.Рез. нуклеиновых кислот
.47
:D506
–D515
.Bernatzky
R
,Tanksley
SD.
1986
.Генетика связанных с актином последовательностей томатов
.Теория Appl Genet
.72
:314
–321
.Bertioli
DJ
DJ
,Cannon
SB
,Broenicke
L
,Huang
G
,Farmer
AD
et al.2016
.Геномные последовательности Arachis duranensis и Arachis ipaensis , диплоидных предков культивируемого арахиса
.Нат Жене
.48
:438
–446
.BUTI
M
,Giordani
T
,Cattonaro
F
,COSSU
RM
,Pistelli
L
, et al.
2011
.Временная динамика эволюции генома подсолнечника, выявленная путем секвенирования и аннотирования трех крупных геномных областей
.Теория Appl Genet
.123
:779
–791
.Camacho
C
,Coulouris
G
,Avagyan
V
,MA
N
,Papadopoulos
J
, et al.2009
.BLAST+: архитектура и приложения
.БМС Биоинформ
.10
.CAVALLINI
A
,Natali
L
,Zuccolo
A
,Giordani
T
,Jurman
I
, et al.2010
.Анализ транспозонов и состав повторов генома подсолнечника ( Helianthus annuus L.)
.Теория Appl Genet
.120
:491
–508
.Чен
ZJ.
2007
.Генетические и эпигенетические механизмы экспрессии генов и фенотипической изменчивости полиплоидов растений
.
Annu Rev Plant Biol
.58
:377
–406
.Dai
S-L
,Wang
W-K
,Li
M-X
,Xu
Y-X.
2005
.Филогенетическое родство Dendranthema (DC.) Des Moul. Выявлен флуоресцентной гибридизацией in situ
.J Интеграт Биол Растение
.47
:783
–791
.De CoSter
W
,D’Hert
S
,Schultz
DT
,Cruts
M
,VAN Brockhoven
C.
2018
.NanoPack: визуализация и обработка длинных данных секвенирования
.Биоинформатика
.34
:2666
–2669
.Доурик
ГДж.
1952
.Хромосомы хризантемы I: вид
.Наследственность (Эдинб)
.6
:365
–375
.EDGER
PP
PP
,PORTEN
TJ
,VANBUREN
R
,CARDIGAN
MA
,COLE
M
, et al.
2019
.Происхождение и эволюция генома октоплоидной земляники
.Нат Жене
.51
:541
–547
.FLYNN
JM
JM
,HUBLEY
R
,GUBERT
C
,ROSEN
J
,CLARK
AG
et et al.2020
.RepeatModeler2 для автоматического геномного обнаружения семейств мобильных элементов
.Proc Natl Acad Sci U S A
.117
:9451
–9457
.Fonseca
N
,Manning
J.
fastq_utils. Гитхаб.GARNATJE
4T
,CANELA
Má
,GARCIA
S
,HIDALGO
O
,PELLICER
J
, et al.2011
.GSAD: размер генома в базе данных Asteraceae
.Цитометрия А
.79
:401
–404
.Джордани
Т
,Каваллини
А
,Натали
Л.
2014
.
Повторяющийся компонент генома подсолнечника
.Curr Plant Biol
.1
:45
–54
.GÖTZ
S
,GARCIAA-GOMEZ
JM
,TEROL
J
,Williams
TD
,Nagaraj
SH
, et al.2008
.Высокопроизводительные функциональные аннотации и интеллектуальный анализ данных с пакетом Blast2GO
.Рез. нуклеиновых кислот
.36
:3420
–3435
.Gurevich
A
,Saveliev
V
,Vyahhi
N
,Tesler
G.
2013
.QUAST: инструмент оценки качества сборки генома
.Биоинформатика
.29
:1072
–1075
.HAAS
BJ
,Salzberg
SLZBERG
SL
,ZHU
W
,PETEA
M
,ALLEN
JE
, et al.2008
.Автоматизированная аннотация структуры эукариотического гена с использованием средства моделирования доказательств и программы для сборки сплайсированных выравниваний
.
Геном Биол
.9
:р 7
.Хемсли
ВБ.
1889
.История Хризантемы
.Гард Хрон
.6
:652
–654
.Hirakawa
Hirakawa
H
,Sumitomo
K
,Hisamatsu
T
,Nagano
S
,Shirasawa
K
, et al.2019
.Сборка полного генома De novo у Chrysanthemum seticuspe, модельного вида хризантем, и ее применение для генетического анализа и анализа открытия генов
.ДНК Res
.26
:195
–203
.Jones
P
,BINNS
D
,Chang
H-Y
,Fraser
M
,LI
W
, et al.2014
.InterProScan 5: классификация функций белков в масштабе генома
.Биоинформатика
.30
:1236
–1240
.Келли
LJ
, ,Лейтч
IJ.
2011
.
Изучение геномов гигантских растений с помощью технологии секвенирования нового поколения
.Хромосомный рез
.19
:939
–953
. 21987187Ким
D
,Сонг
L
,Брайтвизер
FP
,Зальцберг
SL.
2016
.Центрифуга: быстрая и точная классификация метагеномных последовательностей, версия 1.0.4_beta
.биоРксив
.26
:054965
.Кляйн
SJ
,О’Нил
RJ.
2018
.Мобильные элементы: инновации генома, разнообразие хромосом и конфликт центромер
.Хромосомный рез
.26
:5
–23
.Kyriakidou
M
,TAI
HH
,Anglin
NL
,ELLIS
D
,Strömvik
MV.
2018
.Текущие стратегии сборки последовательности генома полиплоидных растений
.Передний завод Sci
.
9
:1660
.Ли
Х.
2013
. Выравнивание прочтений последовательностей, последовательностей клонирования и контигов сборки с помощью BWA-MEM.Ли
Х.
2018
.Minimap2: попарное выравнивание нуклеотидных последовательностей
.Биоинформатика
.34
:3094
–3100
.LI
F
,Вентилятор
G
,LU
C
,Xiao
G
,Zou
C
, et al.2015
.Последовательность генома культивируемого хлопчатника нагорья ( Gossypium hirsutum TM-1) дает представление об эволюции генома
.Нат Биотехнолог
.33
:524
–530
.LIU
B
,Ян
J
,LI
W
,YIN
L
,LI
P
, et al.2020
.Геном Mikania micrantha дает представление о молекулярном механизме быстрого роста
.
Нац Коммуна
.11
:340
.Лю
H
,Ву
S
,Ли
А
,Руан
Дж.
4 0
SMARTdenovo: ассемблер de novo, использующий длинные зашумленные чтения
.Гигабайт
.2021
:1
–9
.Лю
PL
,Ван
Q
,Го
YP
,Ян
J0 9Y
,
2012
.Филогения рода Chrysanthemum L.: свидетельство однокопийного ядерного гена и последовательностей ДНК хлоропластов
.PLoS Один
.7
:e48970
.Лю
Ю.
2020
. Генетическая структура и фенотипические различия среди и внутри существующих популяций Chrysanthemum arcticum L.и С. а. подвид arcticum [магистр наук. Тезис]. Университет Миннесоты.Lukaszewski
AJ
,ALBERTI
,ALBERTI
A
,Sharpe
A
,Киллов
A
,STANCA
ALL
et al.
2014
.Основанная на хромосоме черновая последовательность генома гексаплоидной мягкой пшеницы ( Triticum aestivum )
.Наука
.80
:345
.MA
YP
,YP
,CHEN
мм
,WEI
JX
,ZHAO
L
,LIU
PL
, et al.2016
.Происхождение культурных сортов хризантем — данные ядерных низкокопийных последовательностей гена LFY
.Биохим Сист Экол
.65
:129
–136
.Марсе
G
,Кингсфорд
К.
2011
.Быстрый подход без блокировок для эффективного параллельного подсчета вхождений k-меров
.Биоинформатика
.27
:764
–770
.Nakano
M
,Taniguchi
K
,Masuda
Y
,Kozuka
T
,Aruga
Y
, et al.2019
.Чистая линия, полученная из самосовместимого мутанта Chrysanthemum seticuspe в качестве модельного штамма рода Chrysanthemum
.
Растениевод
.287
:110174
.Natali
L
,COSSU
RM
,Barghini
E
,Giordani
T
,Buti
M
, et al.2013
.Повторяющийся компонент генома подсолнечника, показанный различными процедурами сборки секвенирования следующего поколения, читает
.BMC Genomics
.14
:686
.Нгуен
ТК
,Га
STT
,Лим
ДЖХ.
2020
.Анализ генетического разнообразия хризантем методом генотипирования путем секвенирования
.Hortic Environ Biotechnol
.61
:903
–913
.Оконечников
K
,Conesa
A
,García-Alcalde
F.
2016
.Qualimap 2: расширенный контроль качества с несколькими образцами для данных высокопроизводительного секвенирования
.Биоинформатика
.32
:292
–294
.
Оу
С
,Чен
Дж
,Цзян
Н.
2018
.Оценка качества сборки генома с использованием LTR Assembly Index (LAI)
.Рез. нуклеиновых кислот
.46
:e126
.PENG
Y
,LAI
Z
,Lane
T
,NAGESWARA-RAO
M
,OKADA
M
, et al.2014
.Сборка генома De Novo экономически важного сорняка конского сорняка с использованием интегрированных данных с нескольких платформ секвенирования
.Завод Физиол
.166
:1241
–1254
.PFLUG
JM
,Holmes
VR
,Burrus
C
,Johnston
JS
,MADDISON
DR.
2020
.Измерение размеров генома с использованием глубины считывания, k-меров и проточной цитометрии: методологические сравнения у жуков (Coleoptera)
.G3 (Бетесда)
.10
:3047
–3060
.
Ramírez-Sánchez
O
,Pérez-Rodrígegez
P
,Delaye
L
,Tiessen
A.
2016
.Растительные белки меньше, потому что они кодируются меньшим количеством экзонов, чем животные белки
.Геном Протеом Биоинформ
.14
:357
–370
.Плотва
MJ
,Шмидт
SA
,Борнеман
АР.
2018
.Очистка гаплотигов: переназначение аллельных контигов для сборок диплоидного генома третьего поколения
.БМС Биоинформ
.19
:460
.SCAGLIONE
D
,REYES-CHIN-WO
S
,ACCRADRO
ACHADRA
A
,Broenicke
L
,Portis
E
, et al.2016
.Последовательность генома аутбридинга артишока шаровидного, сконструированная de novo с использованием стратегии низкочастотного секвенирования с учетом фазы потомства F 1
.
Научный представитель
.6
:1
–17
.SCHNable
PS
,WARE
D
,Fulton
RS
,STEIN
JC
,WEI
F
, et al.2009
.Геном кукурузы B73: сложность, разнообразие и динамика
.Наука
.326
:1112
–1115
.Shen
Q
,Чжан
л
,Liao
Z
,Wang
S
,Ян
T
, et al.2018
.Геном Artemisia annua дает представление об эволюции семейства Asteraceae и биосинтезе артемизинина
.Мол Завод
.11
:776
–788
.FA Simão
FA
,Waterhouse
,RM
RM
,Ioannidis
P
,Kriventseva
EV
,Zdobnov
EM.
2015
.BUSCO: оценка сборки генома и полноты аннотации с помощью однокопийных ортологов
.Биоинформатика
.
31
:3210
–3212
.песня
C
,LIU
Y
,песня
A
,DONG
G
,Zhao
H
, et al.2018
.Геном Chrysanthemum nankingense дает представление об эволюции и разнообразии цветков хризантемы и лекарственных свойствах
.Мол Завод
.11
:1482
–1491
.Штапф
О.
1933
.Хризантема макиной
.Магазин ботов Кертиса
.156
.Танака
Р.
1960
.О видообразовании и кариотипах диплоидных и тетраплоидных видов хризантем
.Цитология
.25
:43
–58
.Танака
Р
,Симотомай
Н.
1968
.Цитогенетическое исследование гибрида F1 Chrysanthemum makinoi ×Ch. вульгарный
.Цитология (Токио)
.33
:241
–245
.
Tang
H
,Чжан
x
,MIAO
C
,Zhang
J
,MING
R
, et al.2015
.ALLMAPS: надежное упорядочивание шаблонов на основе нескольких карт
.Геном Биол
.16
:3
.Тодоровская
Е.
2007
.Ретротранспозоны и их роль в эволюции генома растений
.Биотехнология Биотехнология Оборудование
.21
:294
–305
.VAN Dijk
EL
,jaszczzzyn
Y
,Naquin
D
,Thermes
C.
2018
.Третья революция в технологии секвенирования
.Тренды Genet
.34
:666
–681
.VAN GEEST
G
,BOURKE
PM
,VOOOORIPS
,Re
,Marasek-Ciolakowska
A
,Liao
Y
, et al.2017а
.Сверхплотная интегрированная карта сцепления для гексаплоидной хризантемы позволяет проводить мультиаллельный анализ QTL
.
Теория Appl Genet
.130
:2527
–2541
.VAN GEEST
G
,VOOORIPS
Re
,Esselink
D
,Post
A
,Visser
RGF
, et al.2017b
.Убедительные доказательства гексасомного наследования у хризантем, основанные на анализе массива 183 тыс. SNP
.БМС Геном
.18
:1
–12
.Вуртюр
GW
,Sedlazeck
FJ
,Nattestad
M
,underwood
CJ
,Fang
H
, et al.2017
.GenomeScope: быстрое безреференсное профилирование генома на основе коротких чтений
.Биоинформатика
.33
:2202
–2204
.Wang
H
,QI
x
,Gao
R
,Wang
J
,Dong
B
, et al.2014
.Микросателлитный полиморфизм Chrysanthemum sp. полиплоиды: влияние дупликации всего генома
.
Научный представитель
.4
:1
–8
.Warren
RL
,COOBE
L
,Mohamadi
H
,Чжан
J
,Jaquish
B
, et al.2019
.ntEdit: полировка масштабируемой последовательности генома
.Биоинформатика
.35
:4430
–4432
.Фитиль
RR
,Джадд
LM
,Горри
CL
,Холт
KE.
2017
.Unicycler: определение сборок бактериального генома из коротких и длинных ридов секвенирования
.PLoS Comput Biol
.13
:e1005595
.выиграл
SY
,Hwang
Yj
,JUNG
JA
,KIM
JS
,Kang
Sh
, et al.2018а
.Идентификация повторяющихся последовательностей ДНК в геноме Chrysanthemum Boreale
.Sci Hortic (Амстердам)
.
236
:238
–243
.Вон
SY
,Юнг
J-A
,Ким
JS.
2018b
.Полный хлоропластный геном Chrysanthemum Boreale (Asteraceae)
.Ресурс митохондриальной ДНК B
.3
:549
–550
.Вон
SY
,Юнг
J-A
,Ким
JS.
2018c
.Полная последовательность митохондриального генома Chrysanthemum Boreale (Asteraceae)
.Ресурс митохондриальной ДНК B
.3
:529
–530
.выиграл
SY
,Kwon
S-J
,Lee
T-H
,Jung
J-A
,KIM
JS
, et al.2017
.Сравнительный транскриптомный анализ выявляет дупликации всего генома и закономерности отбора генов у культивируемых и диких видов хризантем
.Завод Мол Биол
. 201795
:451
–461
.
Xiao
JOO
J
,Sekhwal
MK
,LI
P
,,Rogupathy
R
,Cloutier
S
, et al.2016
.Псевдогены и их полногеномный прогноз у растений
.Int J Mol Sci
.17
: 27Чжан
H
,ZHU
B
,QI
B
,Gou
x
,Dong
Y
, et al.2014
.Эволюция БАД-компонента мягкой пшеницы на протяжении ее истории на аллогексаплоидном уровне
.Растительная клетка
.26
:2761
–2776
.Примечания автора
© Автор(ы), 2021 г. Опубликовано Oxford University Press от имени Американского общества генетиков.
Это статья в открытом доступе, распространяемая на условиях лицензии Creative Commons Attribution License (https://creativecommons. org/licenses/by/4.0/), что разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы.
org/licenses/by/4.0/), что разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы.Old Japanese Imari Bowl Chrysanthemum Mk #1530949
ПОЛОЖЕНИЯ И УСЛОВИЯ ДЛЯ ПОКУПАТЕЛЕЙ НА ЖИВЫХ АУКЦИОНАХ GOANTIQUES ), должны принять следующие условия для участия в живых аукционах GoAntiques.Покупатель признает, что, принимая настоящие Положения и условия, вы заключаете юридически обязывающее соглашение и соглашаетесь на получение электронных писем и уведомлений об этой продаже, а также размещая ставку во время живого аукциона GoAntiques, что такая ставка означает принятие следующих условий:
1. Премия покупателя: Премия покупателя добавляется к каждой покупке для достижения общей цены продажи. Премия покупателя публикуется на странице сведений о каждом товаре.
2. Общая цена продажи: Общая цена продажи товара включает (i) цену товара с молотка (ii) премию покупателя (iii) применимые налоги с продаж и другие налоги, причитающиеся в связи с продажей, (iv) упаковку , расходы на транспортировку и страхование, а также таможенные сборы, сборы и пошлины, если применимо, и (v) любые другие суммы, причитающиеся с Покупателя, как указано в Политике электронной торговли Продавца, опубликованной на Странице сведений о товаре. С Покупателя может взиматься налог в соответствии с законами и правилами штата, в котором Продавец имеет нексис. Обратите внимание, что продавец собирает премию покупателя и переводит ее в GoAntiques.
С Покупателя может взиматься налог в соответствии с законами и правилами штата, в котором Продавец имеет нексис. Обратите внимание, что продавец собирает премию покупателя и переводит ее в GoAntiques.
3. Онлайн-торги. Регистрируясь для участия в торгах через службу онлайн-аукционов GoAntiques, вы соглашаетесь принять политику GoAntiques в отношении раскрытия истории торгов, включая политику, согласно которой приросты участников онлайн-торгов не сообщаются при торгах против заочной ставки до тех пор, пока не будет сделана заочная ставка. превосходит.
Участники торгов могут оставлять заочные ставки на предметы, которые они хотят приобрести. Для ставок, размещенных на веб-сайте goantiques.com, аукционист GoAntiques будет делать ставки для заочного участника, который будет отображаться во время живого аукциона и в истории ставок ebay как «ставка аукционного зала». Аукционист также будет нести ответственность за защиту резерва продавца, который также будет отображаться как «ставка на аукционной площадке». «Ставка аукционного зала» — это обозначение, которое eBayLiveAuctions дает ставкам, сделанным аукционистом в любой ситуации.Все ставки, оставленные участниками торгов на eBay, будут помечены идентификатором eBay пользователя.
«Ставка аукционного зала» — это обозначение, которое eBayLiveAuctions дает ставкам, сделанным аукционистом в любой ситуации.Все ставки, оставленные участниками торгов на eBay, будут помечены идентификатором eBay пользователя.
Наши тихие аукционы позволяют вам оставлять заочные ставки на любой предмет, на который вы хотите сделать ставку. Начало аукциона в 23:00. и только предметы, которые получили ставки до аукциона, появятся во время аукциона. Если вы видите что-то, что обязательно должно быть у вас, обязательно оставьте свою ставку до начала аукциона.
Пожалуйста, зарегистрируйтесь, чтобы делать ставки заблаговременно до начала. Ни GoAntiques, ни его поставщики или аффилированные лица не несут ответственности за ошибки или невыполнение заявок.Заочные ставки должны быть представлены в электронном виде в течение периода предварительного просмотра на http://www.GoAntiques.com, http://www.ebayliveauctions.com или http://www.eBay.com.
4. Способы оплаты: Приемлемые формы оплаты указаны в описании каждого товара. Способы оплаты различаются для каждого товара в соответствии с Политикой оплаты Продавца.
Способы оплаты различаются для каждого товара в соответствии с Политикой оплаты Продавца.
5. Срок оплаты: Покупатель должен связаться с Продавцом в течение трех (3) дней после завершения аукциона. Полная оплата должна быть произведена после получения уведомления об успешной ставке и должна быть получена Продавцом не позднее, чем через 10 дней после аукциона.Право собственности переходит к Покупателю после полной оплаты. Все расходы по доставке несет Покупатель, если иное не согласовано с Продавцом. Любой риск потери во время доставки будет нести Покупатель после доставки приобретенного товара назначенному общему перевозчику.
Неперечисление платежа Продавцу в срок, указанный в настоящем соглашении, может привести к судебному иску против Покупателя.
6. Выигравшая ставка: Покупателем становится участник, предложивший самую высокую цену, и в случае возникновения спора между двумя или более участниками аукциона координатор аукциона принимает решение о покупателе.
7. Запись о продаже: запись о продаже, размещенная на GoAntiques Live Auctions, будет считаться абсолютной и окончательной во всех спорах.
Запись о продаже: запись о продаже, размещенная на GoAntiques Live Auctions, будет считаться абсолютной и окончательной во всех спорах.
8. Резервная цена: Все лоты выставляются с предварительным резервом. Резерв – это конфиденциальная минимальная цена, ниже которой такой лот продаваться не будет. GoAntiques будет действовать для защиты заповедника, участвуя в аукционе. Аукционист может открыть торги по любому лоту ниже резерва, сделав ставку от имени Продавца, и может продолжать делать ставки от имени Продавца до суммы резерва.Это может быть сделано путем размещения заочных ставок, последовательных ставок или путем размещения ставок в ответ на других участников торгов.
9. Запросы: Все запросы относительно лотов должны быть отправлены непосредственно Продавцу по электронной почте и получены за 24 часа до начала аукциона.
10. Отзыв до продажи: GoAntiques оставляет за собой право изъять любое имущество до продажи и не несет никакой ответственности за такой отзыв
11. Условия гарантии: Все предметы в каталоге продаются, как описано Продавцом пункт.Никакое заявление относительно состояния, вида, стоимости или качества лота не может быть истолковано как явная или подразумеваемая гарантия, заявление или принятие на себя ответственности GoAntiques. Если предмет существенно отличается от описанного Продавцом, у Покупателя есть 48 часов с даты получения имущества, чтобы уведомить Продавца. По истечении 48 часов все предметы считаются «как есть», без права регресса. Возврат наличными или в кредит осуществляется в соответствии с Политикой возврата Продавца.
Условия гарантии: Все предметы в каталоге продаются, как описано Продавцом пункт.Никакое заявление относительно состояния, вида, стоимости или качества лота не может быть истолковано как явная или подразумеваемая гарантия, заявление или принятие на себя ответственности GoAntiques. Если предмет существенно отличается от описанного Продавцом, у Покупателя есть 48 часов с даты получения имущества, чтобы уведомить Продавца. По истечении 48 часов все предметы считаются «как есть», без права регресса. Возврат наличными или в кредит осуществляется в соответствии с Политикой возврата Продавца.
Вышеупомянутая гарантия действительна только для первоначального покупателя.Предметы должны быть возвращены в том же состоянии, что и при покупке. Расхождения в описании решаются взаимно согласованными экспертами, решение которых должно быть оформлено в письменной форме и оплачивается Покупателем.
12. ОГРАНИЧЕНИЕ ОТВЕТСТВЕННОСТИ. GOANTIQUES И ЕЕ ПОСТАВЩИКИ НЕ НЕСУТ ОТВЕТСТВЕННОСТИ ЗА ОСОБЫЕ, СЛУЧАЙНЫЕ ИЛИ КОСВЕННЫЕ УБЫТКИ (В ЛЮБОМ СЛУЧАЕ ВОЗНИКШИЕ, ВКЛЮЧАЯ НЕБРЕЖНОСТЬ), ВЫТЕКАЮЩИЕ ИЗ НАСТОЯЩЕГО СОГЛАШЕНИЯ ИЛИ В СВЯЗИ С ним. НИ ПРИ КАКИХ ОБСТОЯТЕЛЬСТВАХ GOANTIQUES ИЛИ ПАРТНЕРЫ GOANTIQUES НЕ НЕСУТ ОТВЕТСТВЕННОСТИ ПЕРЕД ПОКУПАТЕЛЕМ ПО ЛЮБЫМ ПРЕТЕНЗИЯМ, СВЯЗАННЫМ С ТЕХНОЛОГИЕЙ ЖИВОГО АУКЦИОНА.ЭТО ВКЛЮЧАЕТ ДОСТАВКУ ЭЛЕКТРОННЫХ СООБЩЕНИЙ ПОКУПАТЕЛЯМ. В СЛУЧАЕ ТЕХНОЛОГИЧЕСКОГО ОТКАЗА ВО ВРЕМЯ АУКЦИОНА GOANTIQUES ОСТАВЛЯЕТ ЗА СОБОЙ ПРАВО ОТЗЫВАТЬ СОМНИТЕЛЬНЫЕ ТОВАРЫ И ПОДАТЬ ЗАЯВКУ НА ВЗАИМНЫЙ ОТКАЗ КАК ДЛЯ УЧАСТНИКА, ТАК И ДЛЯ ПРОДАВЦА.
НИ ПРИ КАКИХ ОБСТОЯТЕЛЬСТВАХ GOANTIQUES ИЛИ ПАРТНЕРЫ GOANTIQUES НЕ НЕСУТ ОТВЕТСТВЕННОСТИ ПЕРЕД ПОКУПАТЕЛЕМ ПО ЛЮБЫМ ПРЕТЕНЗИЯМ, СВЯЗАННЫМ С ТЕХНОЛОГИЕЙ ЖИВОГО АУКЦИОНА.ЭТО ВКЛЮЧАЕТ ДОСТАВКУ ЭЛЕКТРОННЫХ СООБЩЕНИЙ ПОКУПАТЕЛЯМ. В СЛУЧАЕ ТЕХНОЛОГИЧЕСКОГО ОТКАЗА ВО ВРЕМЯ АУКЦИОНА GOANTIQUES ОСТАВЛЯЕТ ЗА СОБОЙ ПРАВО ОТЗЫВАТЬ СОМНИТЕЛЬНЫЕ ТОВАРЫ И ПОДАТЬ ЗАЯВКУ НА ВЗАИМНЫЙ ОТКАЗ КАК ДЛЯ УЧАСТНИКА, ТАК И ДЛЯ ПРОДАВЦА.
13. Полное соглашение; Отказ. Настоящее Соглашение, включая Пользовательское соглашение GoAntiques и все экспонаты и другие документы, включенные посредством ссылки, излагают полное понимание и соглашение сторон и заменяют собой любые и все другие письменные или устные соглашения или договоренности между сторонами в отношении предмета настоящего Соглашения. это соглашение.В случае несоответствия между настоящим Соглашением и любым другим документом, являющимся частью настоящего Соглашения, условия настоящего Соглашения имеют преимущественную силу. Время от времени в течение срока действия настоящего Соглашения GoAntiques может вносить поправки в настоящее Соглашение (включая любые включенные документы), публикуя измененные условия на Сайте GoAntiques, отправляя уведомление об изменении Покупателю по электронной почте или иным образом сообщая Покупателю о таких изменениях. Каждая поправка вступает в силу сразу после того, как она первоначально размещена на сайте GoAntiques, отправлена Покупателю по электронной почте или иным образом сообщена Покупателю.Отказ от ответственности за нарушение какого-либо положения настоящего Соглашения не будет действовать и не будет толковаться как отказ от ответственности за любое другое или последующее нарушение. Законы штата Огайо регулируют все сделки.
Каждая поправка вступает в силу сразу после того, как она первоначально размещена на сайте GoAntiques, отправлена Покупателю по электронной почте или иным образом сообщена Покупателю.Отказ от ответственности за нарушение какого-либо положения настоящего Соглашения не будет действовать и не будет толковаться как отказ от ответственности за любое другое или последующее нарушение. Законы штата Огайо регулируют все сделки.
Chrysanthemum indicum L. и C. morifolium Ramat
Действия
‘) var buybox = document.querySelector(«[data-id=id_»+ метка времени +»]»).родительский узел ;[].slice.call(buybox.querySelectorAll(«.вариант-покупки»)).forEach(initCollapsibles) функция initCollapsibles(подписка, индекс) { var toggle = подписка. querySelector(«.цена-варианта-покупки»)
подписка.classList.remove(«расширенный»)
var form = подписка.querySelector(«.форма-варианта-покупки») если (форма) {
var formAction = форма.получить атрибут («действие»)
form.setAttribute(«действие», formAction.replace(«/checkout», «/cart»))
document.querySelector(«#ecommerce-scripts»).addEventListener(«load», bindModal(form, formAction, timestamp, index), false)
} var priceInfo = подписка.querySelector(«.Информация о цене»)
var PurchaseOption = toggle.parentElement если (переключить && форма && priceInfo) {
переключать.setAttribute(«роль», «кнопка»)
toggle.setAttribute(«tabindex», «0») toggle.addEventListener («щелчок», функция (событие) {
var expand = toggle.
querySelector(«.цена-варианта-покупки»)
подписка.classList.remove(«расширенный»)
var form = подписка.querySelector(«.форма-варианта-покупки») если (форма) {
var formAction = форма.получить атрибут («действие»)
form.setAttribute(«действие», formAction.replace(«/checkout», «/cart»))
document.querySelector(«#ecommerce-scripts»).addEventListener(«load», bindModal(form, formAction, timestamp, index), false)
} var priceInfo = подписка.querySelector(«.Информация о цене»)
var PurchaseOption = toggle.parentElement если (переключить && форма && priceInfo) {
переключать.setAttribute(«роль», «кнопка»)
toggle.setAttribute(«tabindex», «0») toggle.addEventListener («щелчок», функция (событие) {
var expand = toggle. getAttribute(«aria-expanded») === «true» || ложный
toggle.setAttribute(«aria-expanded», !expanded)
form.hidden = расширенный
если (! расширено) {
покупкаВариант.classList.add («расширенный»)
} еще {
покупкаOption.classList.remove(«расширенный»)
}
priceInfo.hidden = расширенный
}, ложный)
}
} функция bindModal (форма, formAction, метка времени, индекс) {
var weHasBrowserSupport = window.fetch && Array.from функция возврата () {
var Buybox = EcommScripts ? EcommScripts.Ящик для покупок: ноль
var Modal = EcommScripts ? EcommScripts.Modal : ноль if (weHasBrowserSupport && Buybox && Modal) {
var modalID = «ecomm-modal_» + метка времени + «_» + индекс var modal = новый модальный (modalID)
modal.
getAttribute(«aria-expanded») === «true» || ложный
toggle.setAttribute(«aria-expanded», !expanded)
form.hidden = расширенный
если (! расширено) {
покупкаВариант.classList.add («расширенный»)
} еще {
покупкаOption.classList.remove(«расширенный»)
}
priceInfo.hidden = расширенный
}, ложный)
}
} функция bindModal (форма, formAction, метка времени, индекс) {
var weHasBrowserSupport = window.fetch && Array.from функция возврата () {
var Buybox = EcommScripts ? EcommScripts.Ящик для покупок: ноль
var Modal = EcommScripts ? EcommScripts.Modal : ноль if (weHasBrowserSupport && Buybox && Modal) {
var modalID = «ecomm-modal_» + метка времени + «_» + индекс var modal = новый модальный (modalID)
modal. domEl.addEventListener («закрыть», закрыть)
функция закрыть () {
форма.querySelector(«кнопка[тип=отправить]»).фокус()
} форма.setAttribute(
«действие»,
formAction.replace(«/checkout», «/cart?messageOnly=1»)
) form.addEventListener(
«Отправить»,
Buybox.interceptFormSubmit(
Буйбокс.fetchFormAction(окно.fetch),
Buybox.triggerModalAfterAddToCartSuccess(модальный),
консоль.лог,
),
ложный
) document.
domEl.addEventListener («закрыть», закрыть)
функция закрыть () {
форма.querySelector(«кнопка[тип=отправить]»).фокус()
} форма.setAttribute(
«действие»,
formAction.replace(«/checkout», «/cart?messageOnly=1»)
) form.addEventListener(
«Отправить»,
Buybox.interceptFormSubmit(
Буйбокс.fetchFormAction(окно.fetch),
Buybox.triggerModalAfterAddToCartSuccess(модальный),
консоль.лог,
),
ложный
) document. body.appendChild(modal.domEl)
}
}
} функция initKeyControls() {
документ.addEventListener(«keydown», функция (событие) {
if (document.activeElement.classList.contains(«цена-варианта-покупки») && (event.code === «Пробел» || event.code === «Enter»)) {
если (document.activeElement) {
событие.preventDefault()
документ.activeElement.click()
}
}
}, ложный)
} функция InitialStateOpen() {
var buyboxWidth = buybox.смещениеШирина
;[].slice.call(buybox.querySelectorAll(«.опция покупки»)).forEach(функция (опция, индекс) {
var toggle = option.querySelector(«.
body.appendChild(modal.domEl)
}
}
} функция initKeyControls() {
документ.addEventListener(«keydown», функция (событие) {
if (document.activeElement.classList.contains(«цена-варианта-покупки») && (event.code === «Пробел» || event.code === «Enter»)) {
если (document.activeElement) {
событие.preventDefault()
документ.activeElement.click()
}
}
}, ложный)
} функция InitialStateOpen() {
var buyboxWidth = buybox.смещениеШирина
;[].slice.call(buybox.querySelectorAll(«.опция покупки»)).forEach(функция (опция, индекс) {
var toggle = option.querySelector(«. цена-варианта-покупки»)
var form = option.querySelector(«.форма-варианта-покупки»)
var priceInfo = option.querySelector(«.Информация о цене»)
если (buyboxWidth > 480) {
переключить.щелчок()
} еще {
если (индекс === 0) {
переключать.щелчок()
} еще {
toggle.setAttribute («ария-расширенная», «ложь»)
form.hidden = «скрытый»
priceInfo.hidden = «скрытый»
}
}
})
} начальное состояниеОткрыть() если (window.buyboxInitialized) вернуть
window.buyboxInitialized = истина initKeyControls()
})() .
цена-варианта-покупки»)
var form = option.querySelector(«.форма-варианта-покупки»)
var priceInfo = option.querySelector(«.Информация о цене»)
если (buyboxWidth > 480) {
переключить.щелчок()
} еще {
если (индекс === 0) {
переключать.щелчок()
} еще {
toggle.setAttribute («ария-расширенная», «ложь»)
form.hidden = «скрытый»
priceInfo.hidden = «скрытый»
}
}
})
} начальное состояниеОткрыть() если (window.buyboxInitialized) вернуть
window.buyboxInitialized = истина initKeyControls()
})() .